1. Introduction
Cyanobacteria is an important group of micro-organisms to be studied due to their nutritional simplicity, ubiquitous distribution and in some species, capacity of nitrogen fixation and toxin production.
Higher variation in morphological features among cyanobacteria even within the same species highlights the essential necessity of the contribution of morphological features in identification [1]. Branching patterns, formation of special structures such as heterocysts, akinetes, hormogonia and patterns of motility are the mostly considering properties of interest [1]. Earlier cyanobacteria identification and classification were therefore mainly based on phenotypic and ecological data [2]. However, the ability of adjusting cyanobacteria to fluctuating environmental conditions with morphological and biochemical modifications increases their diversity even at the intra-species level. On the other hand, this morphological and biochemical complexity makes the identification and evaluation of their diversity more and more complicated. Therefore, morphological identification is not sufficient to make conclusions on the classification and the taxonomical state of a particular cyanobacteria strain. Higher variability of morphological features with fluctuating environmental conditions is the major problem of using it alone as the major criterion of cyanobacteria classification [3]. Therefore, molecular characterization is essential for the confirmation of the identity and their state in taxonomy.
Precise identification of cyanobacteria is thus essential in an accurate evaluation of cyanobacteria diversity. Identification of cyanobacteria which is solely based on the morphology would not be reliable as the morphological features are not static as they are modified with environmental fluctuations. This highlights the importance of using a polyphasic approach in the diversity evaluation. A combination of morphological and molecular characterization can therefore be used as a polyphasic approach to evaluate the cyanobacteria diversity.
Sri Lanka is one of the major biodiversity rich representatives in the tropical region with different climatic zones in which many different ecosystems and vegetations are found. There is a huge variation of floral, faunal and microbial diversity among these zones due to remarkable variations in environmental conditions. This tropical continental island bears number of inland freshwater bodies including 103 rivers, over 250 water supply systems [4] and about 10,000 man-made lakes that periodically experience extreme dry conditions which lead to eutrophic conditions [5]. Favorable environmental conditions such as the atmospheric, water and soil temperature, humidity, solar irradiation, pH and precipitation allow not only animals and plants but also mostly undiscovered microbial communities to flourish. Diverse collection of ecosystems and water sources therefore implies the remarkable diversity of cyanobacteria communities among all other microbial communities in this region.
A broad range of fluctuating environmental conditions throughout the year and the presence of number of different ecosystems and water sources within a small region make Sri Lanka one of the best biodiversity hot spots in the tropical Asian region to study the diversity of cyanobacteria. However, the studies carried out so far, exploring the diversity, distribution and the identification of cyanobacteria in Sri Lanka are scarce. Recently few reports have been published on molecular characterization of cyanobacteria in Sri Lanka. Among them, the studies applying a polyphasic approach are even more limited. The cyanobacteria diversity in different water reservoirs, paddy fields, brackish water and tsunami affected areas in Sri Lanka has been studied using a polyphasic approach including the combination of light microscopy, cultures and the analysis of 16S rRNA gene sequences by Wanigatunge et al. [6]. Sethunge and Manage [4] have identified toxin producing, filter clogging, taste and odor forming nuisance algae and some physiochemical parameters in some selected water bodies in Sri Lanka. Bloom formation of cyanobacteria along with dominance of Microcystis sp. was reported in 17 reservoirs of fresh water bodies in Sri Lanka by Jayatissa et al. [7]. Genetic divergence, phylogenetic relationships and potential toxin producing cyanobacteria in reservoirs and well waters in Dry zone of Sri Lanka have been studied using 16S rRNA gene sequences by Liyanage et al. [8]. This study confirms a rich diversity and divergence among toxin producing cyanobacteria in the water bodies in Dry zone in Sri Lanka [8]. Almost all the studies carried out in Sri Lanka so far have been focused on the occurrence of cyanobacteria and their toxic effects. The information on cyanobacterial culturing, biomass harvesting and the use of biomass for various industrial purposes have remained rare or scarce.
Cyanobacteria can provide promising solutions to most of the prevailing global issues. Nutritious food crisis is one such problem. Rapid growth of human population accelerates the depletion of arable lands for cultivation and leads a huge food crisis. Less growth requirements and high nutrient profiles of cyanobacteria make them promising food supplements and have already been used as food sources especially in Asia. Spirulina sp. [9] and Aphanotheca sacrum [10] had been added for their diet by local populations daily basis in some of the Asian countries.
Rapid depletion of protective Ozone layer due to higher amounts of harmful greenhouse gasses has caused harmful Ultra Violet (UV) rays to reach the Earth. Cancerous effects on eyes and skin caused by UV light have thus led to major globally prevailing health issues. With the higher demand for UV protective agents, synthetic UV protective agents were popularized. However, potential toxicity shown by these products has increased the demand for naturally available UV protective agents. Some herbal extracts play a major role in UV protection. Naturally occurring cyanobacteria is one of the best alternatives with high UV protective properties.
Toxin production is one of the well-known natural defense mechanisms in cyanobacteria. Under adverse environmental conditions, some of the strains are producing different types of cyano toxins. Value added potential of C. komarekiana highlights wide industrial applicability of the species thus the safety level of application and consumption should be further studied and confirmed. Therefore, their capability of toxin production and the levels of toxin production should be essentially evaluated before providing recommendations.
Therefore, the main objective of this study was to investigate the diversity of culturable cyanobacteria from freshwater bodies representing three major climatic zones; Dry zone, Intermediate zone and Wet zone in Sri Lanka, using a phylogenetic perspective. The specific objectives were (1) to isolate cyanobacterial strains followed by their morphological characterization (2) to investigate their phylogenetic relationships using the 16S rRNA gene sequence (3) to compare these sequences with other published sequences for deeper taxonomic resolution and (4) to identify and record new species with their value added potential and toxin producing capability.
These goals will improve our knowledge regarding the diversity, ecological distribution and industrial applicability of culturable cyanobacteria strains from Sri Lanka. We hypothesized that freshwater bodies in the tropical region Sri Lanka, harbor an unknown diversity of unicellular and filamentous cyanobacteria that have never been reported for this region.
2. Material and Methods
2.1. Study Area
Sampling was conducted from July 2014 to May 2015. Water samples were collected from freshwater bodies representing Dry zone, Intermediate zone and Wet zone of Sri Lanka including Jaffna peninsula as a major region in Dry zone of Sri Lanka. Sampling was carried out in triplicates, once in each location. Most of the reservoirs selected for sampling were man made and from dry zone of Sri Lanka.
2.2. Sample Collection and Preparation
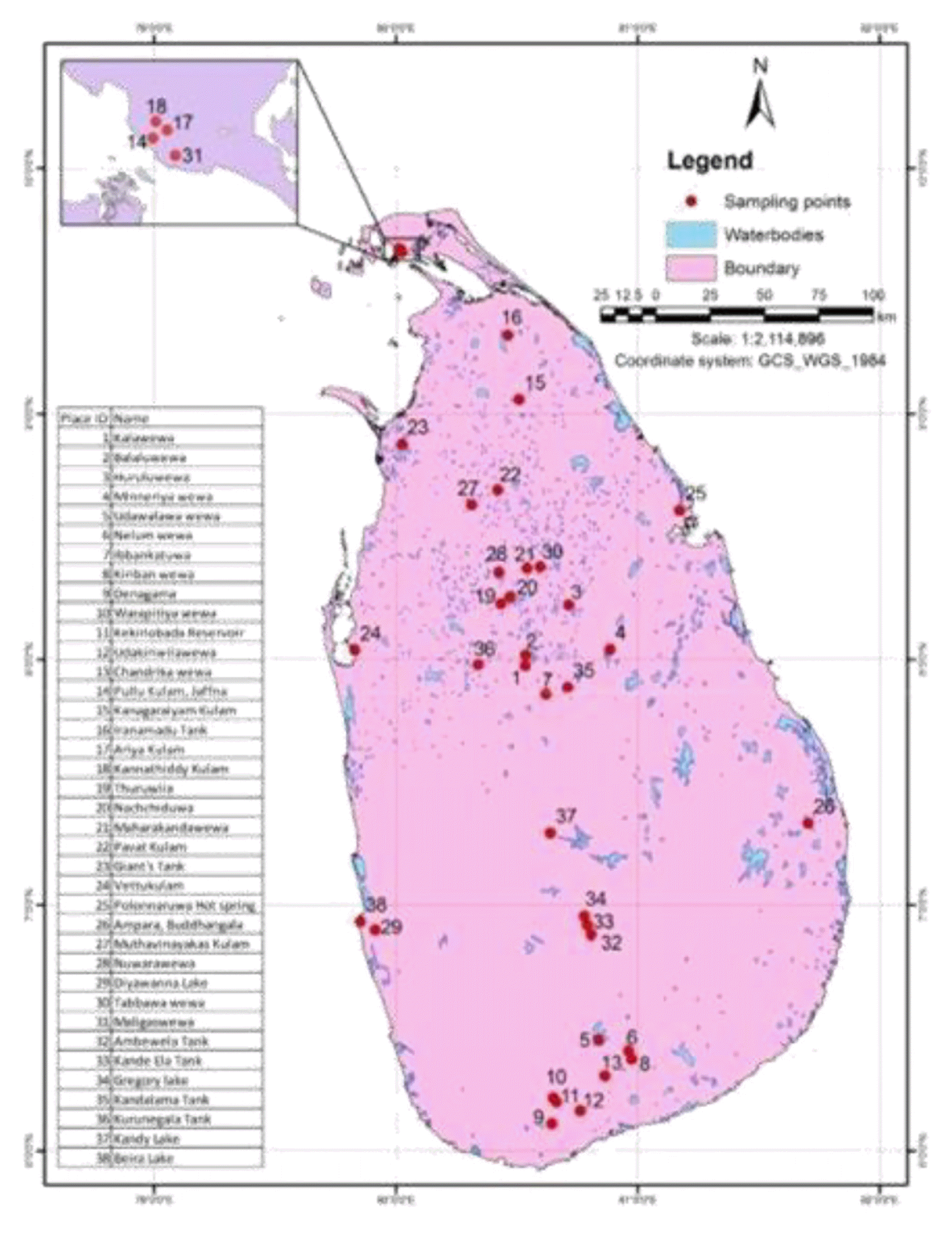
Water was collected from the photic surface layer of the reservoir using a Ruttner sampler and kept in a plastic can till the sample was prepared for culturing (less than two hours). The photic zone of the water was determined by the Secchi disk disappearance technique. The coordinates of all the sampling locations were taken using Geographical Positioning System (GPS) receiver (Garmin, eTrex 30). Sampling locations are summarized in Fig. 1. From each sample 2 L volume was filtered through 20 μm mesh size planktonic net. The retentiate was transferred into a screw cap plastic tube making the final volume into 25 mL by adding distilled water. Another sample was prepared by adding original water to the retentiate instead of distilled water. During sampling, physical parameters (water temperature, pH, and Secchi depth (cm)) of the freshwater bodies in three climatic zones were also measured.
2.3. Cyanobacteria Isolation and Culturing
Ten mL of the planktonic samples were transferred into 50 mL of BG11 [11], GO [12] and ASNIII [13] media for culturing. Ten mL of samples with 40 mL of medium were cultured in 100 mL conical flasks for laboratory cultures and 10 L aspirator bottles with one third filled with media were used for semi mass culturing which were kept at shaking (0 rpm, 200 rpm and 300 rpm) in biological growth chamber under fluorescent light (1000 lux, 1500 lux and 2000 lux light intensity) with constant illumination. Four different pH (7, 7.5, 8, and 8.5) values were selected and growth pH range was optimized based on the average pH range shown by six representative water samples collected from three major climatic zones. The pH of the medium was adjusted using 10% Sodium Hydroxide and 10% Hydrochloric Acid. Number of days taken by cultures in each medium to show their bluish green color growth was recorded. Once growth was observed, 100 μL of samples were sub cultured into petri-dishes by the spread plate method under a laminar air flow cabinet [14] on agar plates containing the same media solidified with 1.5% (w/v) bacteriological agar.
Frequent sub culturing was carried out to isolate single colonies from mixed cultures of cyanobacteria. Repeated streaking and spreading on to the solid medium and microscopic observations were performed until pure monocultures were obtained. Vortexing was used to separate the filamentous cyanobacteria when they were in mixtures. For this, one mL of the sample along with some pieces of glass beads was poured into a micro-centrifuge tube and vortexed. In case of plate culture, algal colonies were picked up using a sharp needle and diluted with water in micro-centrifuge tubes prior to vortexing. Subcultures were obtained by inoculating 100 μL of the vortexed sample on to a petri plate. The purity of the culture was confirmed by repeated sub culturing in plate and regular observation under the microscope [15]. After isolation, all the cyanobacterial strains were cultured and maintained in 250 mL Erlenmeyer flasks filled with 200 mL of BG-11 medium. They were also cryopreserved in a −80°C freezer with cryoprotective agents.
2.5. Molecular Analysis of Selected Cyanobacteria Isolates
Based on the efficiency in biomass production, a total number of 31 isolates were selected for molecular identification with the purpose of evaluating their value added potential for different industrial applications.
2.5.1. DNA extraction and 16S rRNA gene region amplification and sequencing
Total genomic DNA was extracted from the cultured cyanobacterial cells using the method of Smoker and Barnum [18] with slight modifications. The 16S rRNA gene region was PCR amplified in a Techine TC 3000 DNA Thermal Cycler using the primer sets; CYA106F (approx. 675 bp in length) and CYA359F (approx. 450 bp in length) as forward primers and CYA781Ra and CYA781Rb as reverse primers [19].
The cycling profile for the amplification of 16S rRNA gene region included an initial denaturation of template DNA at 94°C for 5 min, followed by 40 cycles of 94°C for 1 min, annealing at 60°C for 1 min, elongation at 72°C for 1 min and a final elongation at 72°C for 15 min. The amplified PCR products were purified using a Wizard SV gel and PCR clean up system PCR purification kit (Promega Corporation, USA) as recommended by the manufacturer. Sequencing of 16S rRNA gene was carried out at Macrogen, South Korea using ABI 3730XL sequencers with amplified samples of respective forward and reverse primers.
2.5.2. BLAST analysis and phylogenetic tree of 16S rRNA gene
The organisms were identified at a molecular level using online Blast software available in NCBI website. The partial sequences obtained in this study were compared with existing sequences for cyanobacteria in NCBI database. The nucleotide sequences generated in this study and the reference sequences retrieved from GenBank were multiple-aligned using CLUSTAL W, Version 1.7. The alignment was corrected manually and converted to a distance matrix. The distance matrix was converted to a phylogenetic tree using the Neighbour-joining (NJ) algorithm [20] of MEGA 5.05; [21] with multiple substitutions corrected and positions with gaps excluded. The 16S rRNA gene sequences determined for the freshwater cyanobacterial strains were deposited in the GenBank database via BankIt submission tool under the accession numbers: KX962074 - KX962104.
2.5.3. Identification and confirmation of the identity of new species C. komarekiana
Depending on the DNA sequencing results, two strains; U08 and U41 were identified as C. komarekiana whose presence in Sri Lanka was identified and reported for the first time. The identity was further confirmed with reproducible DNA sequencing results and morphological characteristics. Furthermore, the value added potential of C. komarekiana was determined as follows.
2.6. Investigation of Value Added Potential of C. komarekiana
2.6.1. Analysis of nutrient content in C. komarekiana
Mass culturing of C. komarekiana was carried out in 100 L size fish tanks under greenhouse environmental conditions with natural light and temperature. Media was prepared at half strength of respective BG-11 at pH 7.5 along with aeration (Risheng RS 2800). Centrifugation, filtering and flocculation techniques (by adjusting pH) were used to harvest the biomass and dry biomass was obtained and made it into a fine powder for further analysis.
2.6.1.1. Total carbohydrate content of C. komarekiana
The Dubois method [22] was used to measure the total carbohydrate content of the fresh dry biomass. The absorbance of the samples was measured at a wavelength of 490 nm using a microplate reader in a spectrophotometer (UV-2450, Shimadzu).
2.6.1.2. Total protein content of C. komarekiana
Lowry method [23] where the absorbance of the sample was measured at a wavelength of 750 nm in spectrophotometer, was used to measure the total protein content of the fresh biomass. The spectrophotometric absorbance was converted to protein concentration using a calibration curve established with Bovin Serum Albumin.
2.6.1.3. Macro and micro mineral contents of C. komarekiana
0.1 g of dried cyanobacteria biomass was measured and placed in the microwave digester and 3 mL of 69% HNO3 was applied and digested them for 25 min. After cooling, the digested volume was transferred to 10 mL volumetric flasks and volumed up to 10 mL with ultra-pure water. Then the diluted volume was filtered using 0.45 μm filter papers. The resulted filtrate was again filtered using 0.45 μm membrane filters and the final filtrate was used for the analysis by using Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES). Among macro minerals, Ca was analyzed by Aggressive Digestion Method using Atomic Absorption Spectrophotometer (AAS)
2.6.2. Measurement of Sun Protection Factor (SPF)
Sun Protection Factor (SPF) was determined by the method described by Dutra et al. [24] followed by the application of Mansur equation shown in Eq. (1) [25]. The absorption data were obtained in the range of 290 to 320, every 5 nm, and 3 determinations were made at each point.
Where: EE – Erythemal effect spectrum; I – Solar intensity spectrum; Abs- Absorbance of sunscreen product; CF – correction factor (= 10). SPF value was determined by UV spectrophotometer [25].
2.7. Determination of Toxin Producing Capability of C. komarekiana
Toxin analysis and quantification were carried out by using ELISA (Enzyme-Linked Immunosorbent Assay) at the Department of Zoology, University of Sri Jayawardenapura, Sri Lanka. Mainly, the toxins; Microcystin-LR and Cylindrospermopsin were analyzed and quantified.
3. Results and Discussion
3.1. Cyanobacteria isolation and morphological identification
In the present study a total of 74 uni-algal cultures were obtained and these were distributed as 52, 16 and 6 from reservoirs of the Dry, Wet and Intermediate zones respectively (Table 1). The morphological characteristics of cyanobacteria which were observed during the present study are summarized in Table S1. Traditionally, the classification of cyanobacteria has been based on morphological characters such as trichome width, cell size, division planes, shape, arrangement, pigmentation and the presence of characters such as gas vacuoles and a sheath [26, 27]. However, some diagnostic features such as gas vacuoles or akinetes can show variations with different environmental or growth conditions and even be lost during cultivation [28, 29]. Such limitations of phenotypic characters have highlighted the requirement for more reliable methods and promoted molecular approaches in cyanobacterial taxonomy including DNA base composition [30, 31], DNA hybridizations [32], gene sequencing [19] and PCR finger-printing [33, 34].
Compared to other climatic zones, Wet zone receives relatively high mean annual rainfall over 2,500 mm without pronounced dry periods. The Dry zone receives a mean annual rainfall of less than 1,750 mm with a distinct dry season from May to September. The Intermediate zone receives a mean annual rainfall between 1,750 to 2,500 mm with a short and less prominent dry season [35]. According to the present study, the highest water temperature (31.35 ± 1.5°C), pH (8.20 ± 0.68) and Secchi depth (75.09 ± 26.31 cm) were recorded in Dry zone whereas all these parameters were recorded the lowest in the Wet zone (Table S2). This could be due to higher rainfall with shorter dry season in the Wet zone which directly influences the water temperature. On the contrary, the lowest rainfall in the Dry zone of the country prolongs the dry season which also influences the water temperature. Favorable environmental conditions such as sufficient amounts of sunlight, water temperature, pH and nutrient rich environment could cause higher diversity of cyanobacteria in Dry zone.
Though 45 genera are reported so far from this tropical region, Sri Lanka [36], a recent study carried out by Wanigatunge et al. [6] reported 26 genera out of them. Another study reports about 40 species of cyanobacteria belonging to 24 genera from Sri Lankan reservoirs [37]. Of these, except for Microcystis aeruginosa, other species are either rare or occur only in small numbers [37]. More recent reports on the occurrence of freshwater cyanobacteria give evidence of Cylindrospermopsis as a dominant form in addition to Microcystis in several reservoirs surveyed [5, 8, 38]. However in the present study Cylindrospermopsis was not observed among the genera isolated and cultured.
3.2. Molecular Identification-The 16S rRNA Region Analysis
3.2.1. Sequencing of 16S rRNA region analysis
The partial sequences obtained in this study were compared with existing sequences for cyanobacteria in NCBI database. All the sequences in the present study showed ≥ 95% similarity with either query coverage or identity or both with existing data. Hence, they were identified with the closest matching data (Table 2). The selection of 31 isolates for molecular identification was based on efficient biomass production.
In the present study, though 74 isolates have been reported based on morphology, molecular identification was carried out only for selected strains which were efficient in biomass production to check their value added potential to be utilized in different industries.
3.2.2. First record on the occurrence of Cephalothrix komarekiana in Sri Lanka
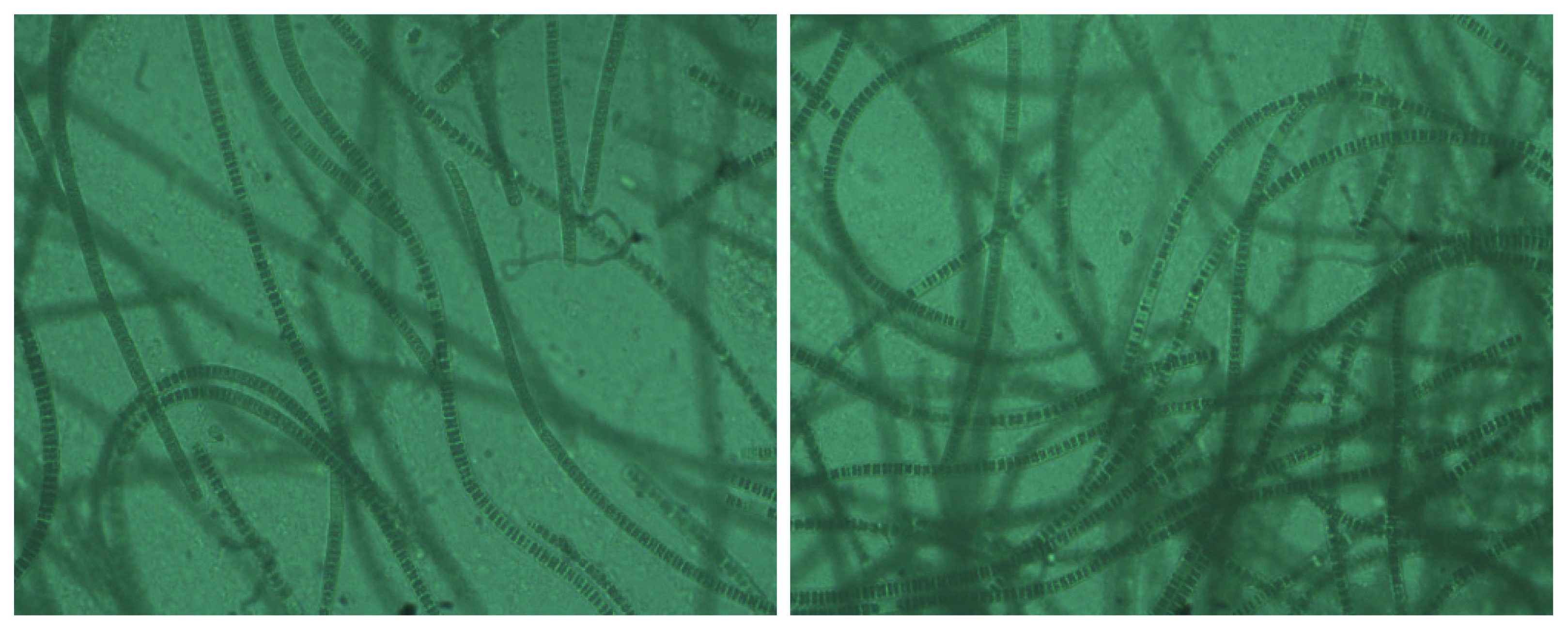
Molecular identification results report the availability of some rare isolates such as Alkalinema pantanalense (U4), Geitlerinema sp. (U36), Westiellopsis prolifica (U58) which were reported in 1986 from this tropical island [36] or rarely been observed thereafter in Sri Lanka. Among them, the novel species Cephalothrix komarekiana was identified for the first time from freshwaters of Sri Lanka (Fig. 2). With morphological and molecular characterization, two isolates; U8 and U41 were identified as Cephalothrix komarekiana. All these two water samples were collected from the freshwater bodies named as Ariya Kulam (U8) and Pollu Kulam (U41) which are located in the northern part of the country, Jaffna peninsula in Sri Lanka. Jaffna peninsula is in the northern most part of Sri Lanka and based on annual average rainfall it covers a major area in Dry zone of Sri Lanka (5° 54′ N - 9° 52′ N and 79° 39′ E - 81° 53′ E). The average mean temperature of the study area varies in between 25.0 to 27.5°C while annual cumulative rainfall is reported in between 1000 to 1500 mm [39]. Rainfall of the study area is seasonal and mainly governed by the North East Monsoon. According to the water quality reports, the freshwater of Jaffna is alkaline in character with high salinity.
3.2.3. Confirmation of the identity of the new species, Cephalothrix komarekiana
Polyphasic approach where the combination of morphological and molecular identification approaches were used, helped in further confirmation of the identity of the new species. DNA sequencing results were highly reproducible to confirm the identity of the isolate U41 as Cephalothrix komarekiana. Both percentage identity and query cover were considered for identification of this strain up to species level. Reproducible results showed 100% query cover and 99.43% percentage identity for U41 (Table 3) confirming the identity of the isolate as Cephalothrix komarekiana.
In previous studies, this novel species was first reported from tropical South American region [40, 41] and a temperate region in Korea [42]. The first report stated that the species was existed in extremely alkaline water and freshwater aquarium in Brazil in 2015 [40] and then from freshwater of Amazonian rivers in the same South American region, Brazil in 2017 [41]. The same species was recorded from brackish waters in Korea in 2018 [42]. However, the identified strains in Sri Lanka were present in freshwater reservoirs used for irrigation purposes. This study has reported the first record of Cephalothrix komarekiana from freshwater reservoirs of Sri Lanka; a country in the Tropical Asia with different environmental conditions.
3.3. Construction of the Phylogenetic Tree with the DNA Sequences Obtained in the Study
According to the constructed phylogenetic tree, it was observed that the isolate U9, U22, U30 and U31 had lower than 95% query coverage or identity. However, the phylogenetic tree with the sequences obtained in the present study for those genera along with previously sequenced sequences obtained from GenBank showed all isolates under the same branch but in different clusters (Fig. 3). Though the isolate U22 was morphologically similar to the two isolates Cephalothrix komarekiana PK41 and Cephalothrix komarekiana AK8, partial sequencing indicated only 91% sequence similarity to the two isolates. By subjecting these three isolates for the total sequencing of the 16S rRNA gene, the confirmation of the species could be achieved.
3.4. Value Added Potential of C. komarekiana
The present study also reports for the first time on the value added potential of Cephalothrix komarekiana present in freshwater bodies in Sri Lanka. The evaluation of value added potential of this newly recorded species will provide a new insight to the applied research and related industrial applications.
3.4.1. Analysis of nutrient contents in C. komarekiana
Total carbohydrate content of the strain of C. komarekiana is 33.23% (Table 4). This value is considerably higher with comparison to the records on proximate available carbohydrate contents present in three microalgae strains; Spirulina plantensis, Chlorella vulgaris and Isochrisis galbana [43]. However, Spirulina plantensis was recorded with a higher average crude protein content of 63.0% [43] whereas the above strain of C. komarekiana showed an average total protein content of 28.93% (Table 4). The fatty acid content of C. komarekiana was recorded in between 12–14% (Table 4) and this strain was among the cyanobacteria strains with considerably higher fatty acid contents. Considerably high major nutrient contents of C. komarekiana suggest their suitability as an alternative food supplement. Furthermore, their nutrient profile for important macro and micro mineral contents will provide a broad picture on their effectiveness and suitability in developing this species as a food supplement.
Nutrient profiles of macro and micro nutrients show that C. komarekiana is rich with higher amounts of macro nutrients such as Mg, K and Ca. Among the tested macro nutrients, Ca was the highest that was analyzed by AAS. Mg was the second highest followed by K (Table 5). Ca was reported as a rich mineral in Spirulina sp. which is already popular as a food source in the world [44]. The reported concentration of Ca in Spirulina sp. was in the range of 1,300–14,000 ppm and it was comparable to the quantity found in milk [44]. Similarly, in this study, the Ca content in C. komarekiana was reported as 4258 ppm. Therefore, C. komarekiana can be identified as a rich source of Ca. As well as Spirulina sp., C. komarekiana therefore becomes a suitable nutrient supplement for vegetarians. Compared to other macro elements, Ca is required by the cellular systems in large amounts as they are essential for cellular communication and modulation including cell movement, muscle contraction, nerve transmission, glandular secretion, blood clotting and cell division. As a Ca rich cyanobacterium, C. komarekiana shows its value added potential to be developed as a low cost, alternative supplement for Ca. Not only Ca but also higher amounts of Mg and K in C. komarekiana suggest its potential to be utilized in food industry and in different medical applications. Iron was in the greatest quantity followed by Mn, Zn, Cu, Co and Ni among all micro nutrients (Table 5). C. komarekiana showed a higher Cu content (4.5 ppm) compared to Spirulina sp. (1.9 ppm) [45]. Mn content present in C. komarekiana (30.3 ppm) is similar to the Mn content in Spirulina sp. (31.2 ppm) [45]. However, Zn (5.7 ppm) and Mg (693.0 ppm) contents in C. komarekiana were lower compared to Spirulina sp. (32.0 ppm and 2120.0 ppm respectively) [45]. Heavy metal quantities of Cd, Cr, Pb, As and Sr in C. komarekiana were in the safe consumption level recommended by the World Health Organization. These concentrations in C. komarekiana were also reported in negligible amounts or in very low concentrations and always lower than the reported levels in Spirulina sp. [45]. It further confirms their level of safety in utilizing them as mineral supplements in food and pharmaceutical industries. As a mineral rich cyanobacterium, C. komarekiana would be another promising natural alternative which can be developed as a food supplement for the improvement of both global public health and economy.
3.4.2. Measurement of Sun Protection Factor (SPF) in C. komarekiana
C. komarekiana: SPF value: 2.37 ± 0.755
According to the above results, the reported SPF value for C. komarekiana is 2.37± 0.755. Compared to the SPF values obtained for some of the herbal extracts namely; Aloe vera, Carrot, Cucumber and Watermelon by Malsawmtluangi et al. [46], C. komarekiana has shown considerably higher SPF value. Therefore, it suggests the effectiveness of C. komarekiana as a promising botanical agent to be utilized and developed as a UV protective product in cosmetic industry. Higher photosynthetic ability and growth rate, less nutrient and land area requirement, capability of mass production with less capital investment and most importantly zero environment pollution further highlight the value and effectiveness of utilizing this botanical agent over other synthetic products and plant extracts.
Suitable nutrient profile and efficient and high UV protective properties suggest the value added potential of C. komarekiana and provide a clear insight on possible industrial applications. However, some cyanobacteria strains are toxin producing. Therefore, recommendation of the species for possible industrial applications is still one step away and requires further confirmation of their level of risk for applications.
3.5. Toxin Analysis of C. komarekiana
Toxin producing capability of C. komarekiana for Microcystin-LR and Cylindrospermopsin was investigated. Total Microcystin-LR for C. komarekiana was 0.63±0.045 μg/L. The detected concentration of total Cylindrospermopsin for the strain was 0.06± 0.085 μg/L. According to the World Health Organization (WHO)’s and the Oregon Health Authority (OHA)’s guidance on cyanotoxins, a provisional guideline value of 1.0 μg/L has been adopted for Microcystin-LR. The above guideline value is applicable for all cell bound and extracellular microcystins [47, 48]. Provisional lifetime guideline value set by WHO for Cylindrospomopsin is 0.7 μg/L and short-term guideline value is 3.0 μg/L [47]. The OHA’s provisional guideline value for Cylindrospomopsin is 1.0 μg/L [48]. All these values have been calculated considering average adult body weight of 60 kg as a standard [47, 48]. With comparison to all these standard values, all the produced toxin concentrations are below the risk level for human health. Therefore, these values further confirm the suitability of using C. komarekiana as a food supplement, effective natural sun protective agent and in other industrial applications.
4. Conclusions
The present study was carried out to investigate the diversity of culturable cyanobacteria from freshwater bodies in the tropical Asian region Sri Lanka, with a phylogenetic perspective. A total of 74 uni-algal cultures were obtained and these were distributed as 52, 16 and 6 from reservoirs of the dry, wet and intermediate zones respectively. Among them, C. komarekiana was recorded for the first time from tropical region; Sri Lanka with the availability of some other rare isolates such as Alkalinema pantanalense, Geitlerinema sp., and Westiellopsis prolifica. Rich nutrient profiles of total carbohydrates, total proteins, fatty acids and macro and micro elements showed its capability to be developed as natural, safe alternative food and mineral supplements. Rich Ca profile in C. komarekiana suggests its potential utilization separately as a Ca supplement which would be useful in treating complications caused by Ca deficiencies. C. komarekiana has also been proved to be an efficient natural UV protective agent. Suitable nutrient profiles and efficient UV protective properties provide a clear insight on a wide range of possible industrial applications of C. komarekiana. Their ability and concentrations of toxin production further confirmed the safety level of utilization of this species for the improvement of public health. Higher photosynthetic ability and growth rate, less nutrient and land area requirement, capability of mass production with less capital investment and most importantly zero environment pollution nominate this species as one of the best natural agents which can be utilized in finding eco-friendly solutions for many of the global issues.