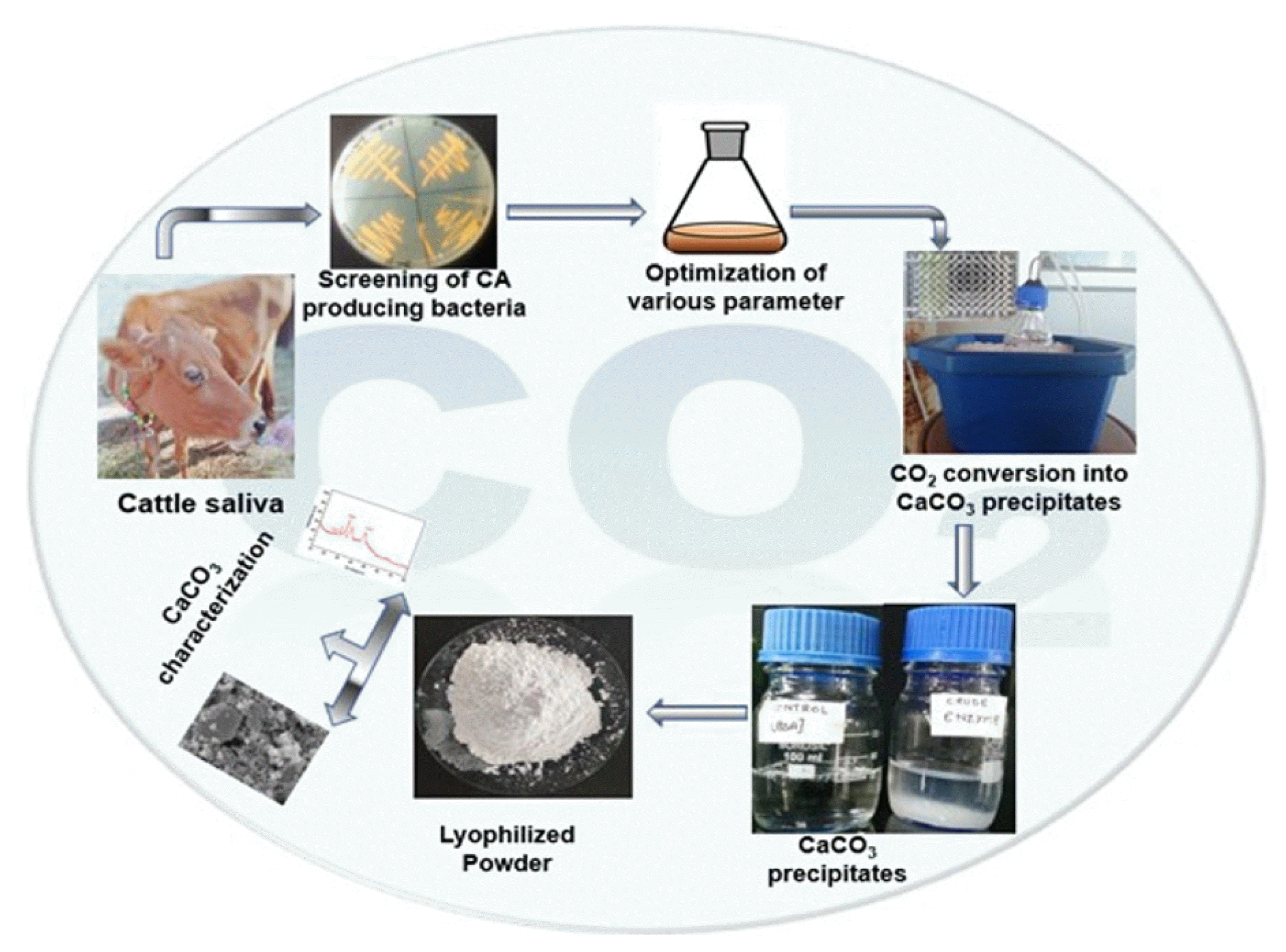
1. Introduction
Global warming is one of the serious issues due to excessive emission of carbon dioxide (CO2) into the atmosphere. To reduce CO2 emission, numerous government and non-governmental organizations endorse to switch from fossil fuel power to clean energy sources to protect the atmosphere from the terrible effects of global warming [1–3]. Furthermore, worldwide research is going on for finding a potent and eco-friendly method that directly converts industrial emitted CO2 into a useful product. Some of the algae, bacteria, and cyanobacteria play a vital role in alleviating the increasing level of CO2 using their carboxylating enzyme [4, 5]. Recently, these carboxylating enzymes are getting attention due to its role in CO2 sequestration [1, 6]. However, algae have good potential for CO2 fixation, but it has a few disadvantages such as light requirement [7]. Hence, it is advantageous to isolate a CO2 fixing bacteria or enzyme which does not require continuous light for its growth and activity [8].
CA is one of the fastest known biocatalysts and involved in the CO2 sequestration. CA contains zinc in its active center and converts CO2 to bicarbonates, which can be further converted to CaCO3 in the presence of calcium ion. The application of CA in the conversion of CO2 from flue gas into thermodynamically stable, environmentally safe calcium carbonate offers several advantages [9]. First of all, the process is extremely fast and take place near ambient condition. CaCO3 formed during CO2 conversion can be utilized in the preparation of white pigment, cement, antacids and others [10]. Thus, a new biomimetic approach using CA has been found to be viable for fixing a huge quantity of CO2 into CaCO3. CA catalyzes several other hydrolytic reactions too, including hydration of urea, carboxylic acid, halides, and hydrolyzable substrates [11, 12]. The CA catalyze the hydration of CO2 in two steps ping-pong mechanism. Its active site has divalent metal ions generally zinc in a tetrahedral confirmation, containing three amino acid as ligands and hydroxide ions coordinating the metal [13]. CA is present in eukaryotes and prokaryotes, where it takes part in various physiological functions such as acid-base balance, hemostasis, respiration, and photosynthetic CO2 fixation [14]. Although CA is present ubiquitously but microbial CA has received more attention due to ease of their production and applications. Till now, CA has been classified into five (α, β, γ, δ and ɛ) different classes [15]. The first studied, class is the α-CA as it plays a crucial role in human pathology and drug targeting. However, the other classes take part in the conversion of gaseous CO2 [16]. The β class exists as a dimer, tetramer and hexamer and mostly present in algae, bacteria, and archaea. The γ class is mostly found in archaea and exists as a trimer [17]. This enzyme is either present extracellularly or inside (intracellular) the cytoplasm in the bacteria [18].
Recent studies suggest that CA is widespread in bacteria and archaea domain, indicates that this enzyme has a more extensive and vital role in prokaryotic physiology [19]. CA producing bacteria were isolated from various sources such as deep sea-water, alkaline soil, mangroves soil, and seashore water [20]. Moreover, the optimization of various parameters is essential to enhance the total yield, maximal activity of the enzyme and to reduce the process cost. The high enzyme titer has been attained by changing the ratio of various media ingredients that are influencing the growth and enzyme production in bacteria. The optimization is greatly influenced by various physicochemical and nutritional factors such as temperature, agitation rate, pH, media components, inoculum volume, etc. Thus, optimizing the best conditions for maximum enzyme production is still vital and essential in biocatalytic transformations.
In the present study, CA producing bacteria was isolated from cow saliva and the parameters were optimized for the production of CA in bulk amount. The application of crude CA has been studied for the conversion of CO2 into calcium carbonates (Fig. 1). As per our knowledge, this is the first report on the isolation of CA producing bacteria from cow saliva. In the current global warming scenario, this work might be helpful in CO2 conversion and environmental amelioration. Here, we demonstrate that isolated bacteria are the green biocatalyst for the conversion of CO2 into calcium carbonates.
2. Methods
2.1. Isolation of a Bacterial Strain
CA is a ubiquitous enzyme present in all plants, mammalian tissues, algae, and bacteria [19]. In the present study, the sample was collected from six years old cow saliva from District Mandi, Himachal Pradesh, India with the help of sterilized bud (Fig. 2). The saliva was serially diluted and then 100 μL of culture was transferred onto nutrient agar having 3mM p-nitrophenyl acetate (p-NPA), incubated at 30°C for 48 h. The appearance of the intense yellow colour colonies indicated the production of carbonic anhydrase [21]. The bacterial isolates, which utilized p-NPA on agar plates, were further screened for CA activity in nutrient broth. For CA activity assay, the seed culture was prepared aseptically in 50 mL nutrient broth containing a loopful culture of T5 isolate and incubated at 30°C for 24 h. For the production of CA, the nutrient broth was used as production media and the seed culture 2% v/v was used as inoculum for 50 mL production medium. The inoculated production medium was incubated at 30°C for 36 h at 120 rpm. The broth was centrifuged at 10,000 rpm for 10 min after 36 h of incubation. Then pellet was suspended in phosphate buffer (pH 7.0) and homogenized well. The suspension was sonicated using 20 kHz frequency for 5 min at 4°C and centrifuged at 10,000 rpm for 20 min. The supernatant was used as crude enzyme extract.
2.2. Enzyme Assay
CA activity was assayed in the culture broth using a previously reported method by measuring the micromole of p-nitrophenol released from p-NPA [22]. To 825 μL of Phosphate buffer (50 mM, pH 7.5), 175 μL of the substrate stock solution (p-NPA, 10 mM in isopropanol) was added. Then, the reaction mixture was incubated in a water bath at 37°C for 5 min. Subsequently to initiate the reaction 25 μL of an enzyme was added. The reaction mixture was again incubated at 37°C in a water bath for 5 min [23]. The amount of p-nitrophenol released was measured at 410 nm after 5-min incubation using a microplate reader (Thermo-scientific, Multiskan™ FC Microplate Photometer) [24]. Based on CA activity, a potent isolate T5 was selected for all subsequent studies. The colonies of T5 isolate was subjected to gram’s staining and observed at 100 X magnification under a compound microscope. The biochemical characterization of T5 isolate was conducted according to Bergey’s Manual of Systemic Bacteriology [25].
2.3. Identification of Bacterial Strain
The bacterial strain was identified by 16S rRNA sequencing by Bioreserve Biotechnologies Pvt. Ltd., Hyderabad, India [26]. The nucleotide sequences obtained from 16s rRNA sequencing were subjected to a homology search using BLAST and aligned using a MEGA X software. Then, the phylogenetic tree was made using the neighbour-joining method.
2.4. Optimization of Production Parameter for CA Producing Bacteria
Luria broth, nutrient broth, peptone broth, minimal salt media, muller Hinton broth, and basal salt media were used for enzyme production. Luria broth, nutrient broth, and muller hinton broth were procured from Hi Media, Mumbai. The composition of other media used for enzyme production was peptone broth (g/L): beef extract 3.0, glucose 1.0, NaCl 5.0, peptone 5.0, CaCO3 6.0; minimal salt media (g/L): KH2PO4 6.8, MgSO4 0.2, Na2HPO4, 7.8, ZnCl2 0.02, ZnSO4.7H2O 0.05, NaNO3, 0.085; and basal slat media (g/L): sucrose 5.0, MgSO4.7H2O 0.5, Na2HPO4 2.0, FeCl3·6H2O 0.005, CaCO3 0.1. The effect of temperature on the CA production was studied by incubating production culture at various temperatures ranging from 20 to 60°C. To determine the effect of inoculum volume (%v/v) on CA production, production media was inoculated with a (2–12 %v/v) culture of T5 isolate and incubated at 40°C. The optimum agitation rate for CA production was determined by incubating the production media at 40°C at varying agitation speeds (80, 100, 120, 140, 160, and 180 rpm). The enzyme activity was determined for each incubated production media sample using standard activity assay [23].
2.5. Optimization of Reaction Parameter for CA Producing Bacteria
For optimizing reaction conditions of CA effect of reaction time (2–18 min), different buffer systems (citrate buffer, potassium phosphate buffer, and Tris–HCl buffer, 50 mM), reaction temperature (25–60°C), different metal ions (Na+, K+, Ca2+, Zn 2+, Mg2+, Fe3+, and Al3+) and organic solvent (ethanol, isopropanol, ethanediol, n-butanol, propandiol, and, acetonitrile) was studied on CA activity.
2.6. Enzymatic Conversion of Carbon Dioxide into Calcite
The potential of CA for CO2 conversion was determined as described by Giri et al. [27], with minor modifications. The precipitation of CO2 into calcium carbonate (CaCO3) was carried out in a 50 mL total reaction mixture containing 2 mL Tris buffer (1M, pH 8), 23 mL of 2 (%v/v) calcium chloride solution, 23 mL of CO2 saturated water and 2 mL (0.5 mg/mL) of an enzyme in phosphate buffer (50 mM, pH 7). The reaction was performed for 10 min at room temperature. Bovine serum albumin (BSA) was used as a control. The precipitate formed after 10 min was recovered by centrifugation. The sediment of precipitates was lyophilized overnight to obtain the dry powder. Then, CaCO3 precipitates were weighed to determine the amount of carbonate deposited during the enzymatic reaction.
2.7. Instrumental Analysis of CaCO3
The CaCO3 precipitates were lyophilized to get dry powder of CaCO3. The CaCO3 precipitates were scanned in the range of 400–4,000 cm_1. Compositions of the precipitated solid crystals were examined by using X-ray diffraction (XRD). To determine crystal morphology Scanning Electron Microscopy (SEM) was performed by using JSM7401F (JEOL).
3. Results and Discussion
3.1. Isolation of CA Producing Bacteria
The ten bacterial isolates from a cow saliva sample, which efficiently utilized p-NPA on agar plates, were selected for secondary screening and cultivated in nutrient broth to detect CA activity. p-NPA plates assay is specific and based on the formation of yellow-colored (p-NP) product formed around the colonies. Among these bacterial isolate T5, showed maximum CA activity, and selected for further work (Fig. 3 (a)). The T5 isolate was rod-shaped and gram-positive bacilli (Fig. 3 (b)). And the results of the biochemical analysis show Voges-Proskauer positive, nitrate reduction positive, glucose and sucrose positive. The presence of CA in mammalian saliva is reported several decades ago. Yoshimura et al. [28] suggested that the role of CA in mammalian saliva is to regulate saliva pH. Furthermore, the screening of microorganisms from cow saliva for CA production is the rapid and cost-effective method. Jaya et al. [29] reported an isolate Bacillus safensis from the water sample that produced CA. Similarly, Kupriyanova et al. [30] reported that the presence of CA in Microcoleus cathonoplastes, isolated from the soda lakes in Russia. Various pathogenic bacteria such as Yersinia pseudotuberculosis and Listeria monocytogenes isolated from soil and water were reported to produce CA [31]. Microorganisms need CO2 and bicarbonate in their metabolic activities and CA plays a vital role to meet these demands.
3.2. Identification of Strain
The genomic DNA was isolated from bacterial isolate T5 and its DNA analysis was done by agarose gel electrophoresis (Fig. 4 (a)). The 16s rRNA was amplified from genomic DNA using universal primers. Then bacterial strain was identified as Corynebacterium flavescens by 16S rRNA sequencing based on phylogenetic analysis and nucleotide homology. The phylogenetic tree was made by using a neighbour tree joining method showing maximum sequence homology of 99.42% and 99.27% with Corynebacterium flavescens HBUM07012 and Corynebacterium flavescens EHFS1S12Hc, respectively (Fig. 4 (b)). The gene accession number of the organism was MN982752 deposited in NCBI.
3.3. Optimization of Production Conditions for CA
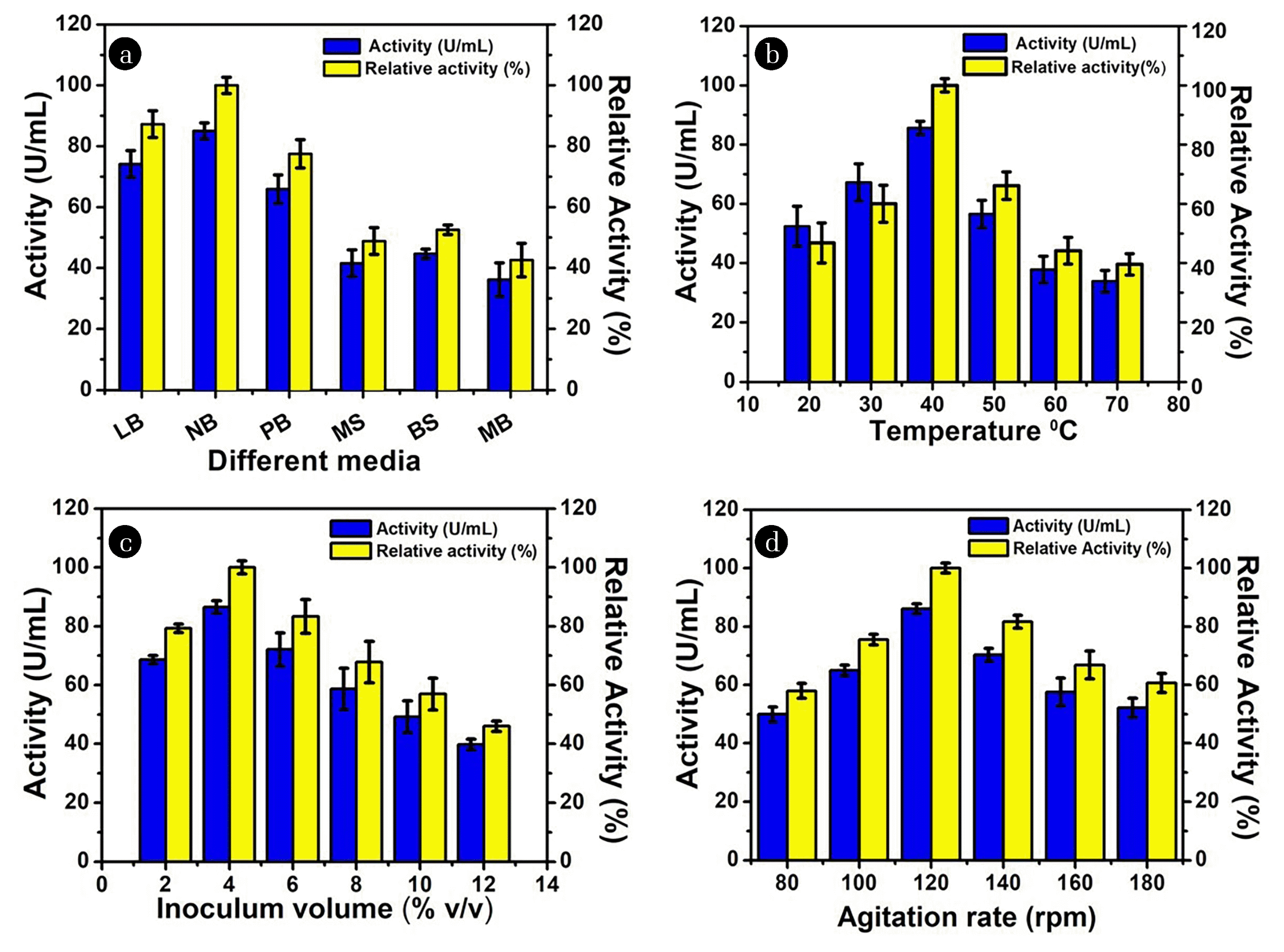
Optimization of production and reaction parameters is widely known to rise the production significantly. In the present study, out of six media, the maximum CA activity (84.99 U/mL) was observed in nutrient broth media (Fig. 5 (a)). In contrast, to the present findings, Sharma et al. [23] reported maximum CA production from Pseudomonas fragi in peptone broth. Also, in a previous study, the nutrient broth was reported as the best medium for the production of CA from Nocardiopsis lucentensis [20]. Thus, the nutrient broth media was selected for further studies. The temperature of incubation significantly affects the production of microbial enzymes and their activity. The maximum production of CA with residual activity 85.59 U/mL was observed at 40°C. Furthermore, a decrease in the CA activity was noticed with a further increase in temperature (Fig. 5 (b)) [24]. The decrease in activity might be attributed to the denaturation of protein at a higher temperature. Similarly, Sharma et al. [23], reported the optimum CA production was observed at 40°C. In a recent study, Bacillus safenis was observed to show optimum activity at 40°C [29]. In another study, CA from Citrobacter freundii was found to be active at 37°C [27]. However, the optimum temperature for CA production from Methanobacterium thermoautotrophicum was found to be 75°C [22]. These results showed that CA production has its own temperature optima and is species-specific which favours maximum CA production.
The volume of inoculum also affected the production and activity of enzymes from the isolated bacterial strain. An increase in CA production was observed with an increase in an inoculum volume up to 4% v/v then it started decreasing (Fig. 5 (c)). Another study showed that 1.5% inoculum size was optimum for CA production from Aeribacillus pallidus [32]. Usually increased inoculum volume improves the growth of bacteria up to a certain level and after that bacterial growth starts decreasing due to nutritional limitation. Furthermore, lower inoculum volume has a low number of bacterial cells in medium and this requires a longer time to grow. This might lead to the accumulation of toxic compounds which ultimately decreased enzyme production [33]. However, some studies reveal that an increase in inoculum size results in more enzyme production and vice versa. The maximum CA activity was observed at an agitation speed of 120 rpm i.e., 86.50 U/mL (Fig. 5 (d)). The CA activity was increased by increasing the agitation rate up to 120 rpm. But the high agitation rate also resulted in decreased activity due to mechanical disruption of proteins. In a previous study, carried out by Zhang et al. [34], showed that 150 rpm most appropriate agitation rate for the production of CA whereas CA from Aeribacillus pallidus gave maximum activity at 200 rpm [32]. The key role of agitation in the fermentation broth is to ensure adequate mixing of oxygen that eventually becomes available for the growth of bacteria in the dissolved form of oxygen. Also, microbial enzyme production is observed to be good under continuous shaking conditions as compared to static conditions [35].
3.4. Optimization of Reaction Parameters for CA
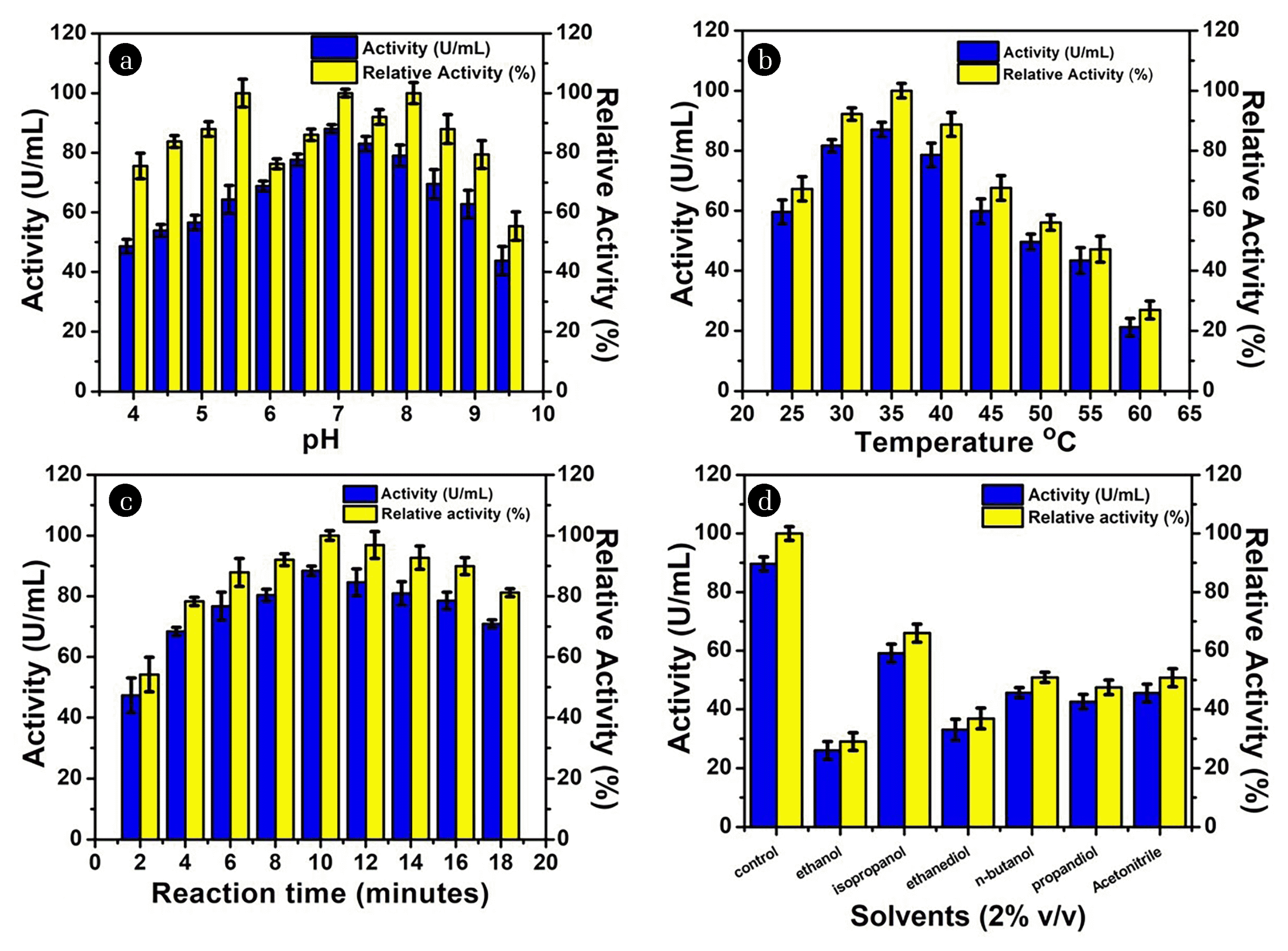
The effect of pH on CA activity was determined and optimum pH for maximum CA activity was 7 in phosphate buffer (Fig. 6 (a)). A decrease in activity at high and low pH generally results in loss of enzyme activity due to disruption of three-dimensional structure and alteration in amino acid residues present in the active center of an enzyme. In another study, CA from Bacillus sp. gave maximum activity at pH 8 whereas CA from Lactobacillus showed optimum activity at pH 6 [36, 37]. However, CA from Helicobacter pylori showed optimum activity in an acidic environment and high acid tolerance [38]. The effect of reaction temperature on the CA activity was studied. The result presented in (Fig. 6 (b)) showed that the optimum temperature for CA activity was 35°C after that decline in CA activity was observed. In the previous study, the optimum temperature for purified CA was found to be 60°C [36]. Mostly, CA isolated from human erythrocyte and bovine shows CA activity in the temperature range of 35–40°C and in the pH range of 6.5–7.5 [39]. The optimum reaction time for CA activity was observed after 10 min of incubation (88.55 U/mL) (Fig. 6 (c)). After 10 min decrease in enzyme activity was observed which may be due to the inhibition of product and denaturation of the enzyme. All these facts indicate the functional diversity of CA and the capability of this enzyme to perform diverse roles in the organisms living in extreme environments.
Out of various organic solvent used, all of them decrease CA activity (Fig. 6 (d)). This can be credited to conformational changes in enzyme active sites that are responsible for the decrease in enzyme activity [40]. Furthermore, the effect of metal ion on CA activity at concentrations 1mM and 5mM was studied (Table 1). The presence of metal ion in the reaction medium either inhibits or promotes the enzymatic reaction. CA activity was found to be increased in presence of Zn2+ and Fe3+ ion this might be due to reason these metal ions are present in the active site of CA, they could have a stabilization effect thus enhancing the CA activity. In contrast, CA activity was not affected by the presence of Ca2+ ion indicates that enzyme could perform in the presence of calcium ion. Whereas, CA activity was inhibited by the presence of Na2+, K+, Mg2+, and Al3+ ions. In a previous study, CA activity from Pseudomonas fragi was significantly enhanced by Zn2+, Fe3+ and Cd2+, Na2+, K+, Mg2+, and Ca2+ had no effect, whereas Hg2+ and Pb2+ had an inhibitory effect [23]. In contrast, the activity of CA from Dicentrarchus labrax was inhibited by Zn2+, Hg 2+, Al3+, Cu2+, and CO2+ [41].
3.5. Enzymatic Conversion of Carbon Dioxide Using Crude Enzyme
Finally, the application of CA to convert CO2 into CaCO3 in the presence of calcium ion was examined. Test reaction containing crude CA showed a significant conversion of CO2 as compared to that of control. BSA was used as a control in the experiment which showed no precipitation of CO2. The CO2 conversion efficacy of CA was studied by calculating the amount of CaCO3 synthesized. The crude CA resulted in the formation of 45 mg CaCO3/mg of protein. The CO2 conversion efficiency of the enzyme produced from Corynebacterium flavescens is much higher as compared to CA from Bacillus pumilis (33.06 mg CaCO3/mg of protein) and Pseudomonas fragi (27.33 mg CaCO3/mg of protein) [42, 23].
CaCO3 is commonly used in various industries such as fillers for paints, papers, plastics, as well as food and pharmaceutical applications [43]. Therefore, it confirmed that the bacterial isolate is producing potent CA which can be used for the CO2 conversion in the lab as well as commercial scale.
3.6. Instrumental Analysis of CaCO3
In the present studies, the crude enzyme precipitates CaCO3 and exhibited morphological resemblance with CaCO3 standard taken from Sigma Aldrich. In the SEM analysis, spherical vaterite crystal was formed in control and rhombohedral vaterite and calcite crystals were formed in the presence of an enzyme (Fig. 7 (a) and (b)). This result from SEM analysis conforms to the finding of Jo et al. [44]. The composition of precipitated powder was confirmed by XRD analysis. The two forms of CaCO3 were vaterite, and calcite, analyzed by XRD patterns. Diffraction peak at 29.38, 34.04, 43.90 corresponds to the calcite crystal phase whereas the peak at 22.54, 27.08 corresponds to the vaterite crystal phase (Fig. 7 (c)). Furthermore, various studies have shown that calcite and vaterite were phase formed in the presence of CA [37, 44].
4. Conclusions
Microorganisms are commonly used to produce the enzymes of industrial utility. The isolation of CA from cow saliva is cost-effective as the price of commercially BCA is about $3,000 g−1. Thus, there is a price limitation for using BCA in various CO2 conversion applications. The improved CA production from Corynebacterium flavescens was achieved by the optimization of various production and reaction parameters. The high yield of CA was achieved from this process as an indication of the possibility for its large-scale production. The crude enzyme was also found to be effective in the CO2 conversion experiment and hence its efficacy in CO2 capture can be exploited at the industrial level too. In future studies, the CA enzyme can be purified and used in an immobilized form for better reaction efficiency. Moreover, directed evolution and rational hybrid design can be used to develop the mutant of this enzyme with better enzyme stability for industrial purposes.