1. Introduction
Sediment microbial fuel cells (SMFCs) are attractive bioelectrochemical technologies that produce electricity from organic matter in sediment via metabolism of exoelectrogens [1, 2]. SMFCs attract increasing attentions as power sources to drive remote sensors to monitor environmental qualities of rivers, lakes and marine with the advantages of low cost maintenance and long-term stability [3–7]. Moreover, SMFCs have caught much attention as an effective technology to in situ remediate contaminated soil/sediment by organic or inorganic matters, via enhanced extracellular electron transfer process through accumulation of exoelectrogens [8–10].
For a typical configuration of SMFCs, the cathode is generally floating in the water layer to accept electron donors such as dissolved oxygen, while the anode is embedded in the sediment to accumulate anaerobic exoelectrogens on the anode surface [11–13]. Previous studies revealed that sediment characteristics, i.e. the concentration of O2, NO3−, SO42−, varied with the depth of the sediment [14, 15], accompanied with different types and physiological characteristics of microorganisms. Therefore, the depth of anode location in SMFCs was verified as an important factor affecting the electric power generation of SMFCs [1, 16]. Exoelectrogens are a key component of SMFCs [17]. Their relative abundance in anodic biofilm and electro-activity affect greatly the performance of SMFCs in terms of electric energy output and contaminants removal. Couples of research analyzed the microbial community structure of anodic biofilm in SMFCs [18, 19], while few research paid attentions to the influences of burial depth of anode on microbial diversity.
Knowledge on microbial communities of anodic biofilms with different burial depth is limited by the use of clone library methods with low sequencing depth [20]. The sequencing analysis was conducted with 454 pyrosequencing to deeply understand the microbial community structure [21–23]. In the present study, the performance of SMFCs with different anode burial depth was recorded and analyzed. The microbial diversity and exoelectrogens of anodic biofilms were revealed with the aid of 454 pyrosequencing to obtain insight into the regulation of anode burial depth on the performance of SMFCs.
2. Material and Methods
2.1. Sediment and Water Samples
Both sediment and water were sampled from a pond in Xiamen city, Fujian province. Sediment was gently separated by hand and passed through a 0.5 cm mesh to remove coarse debris, and then thoroughly mixed using a mechanical stirrer to ensure that the sediment was homogeneous.
2.2. SMFCs Construction and Operation
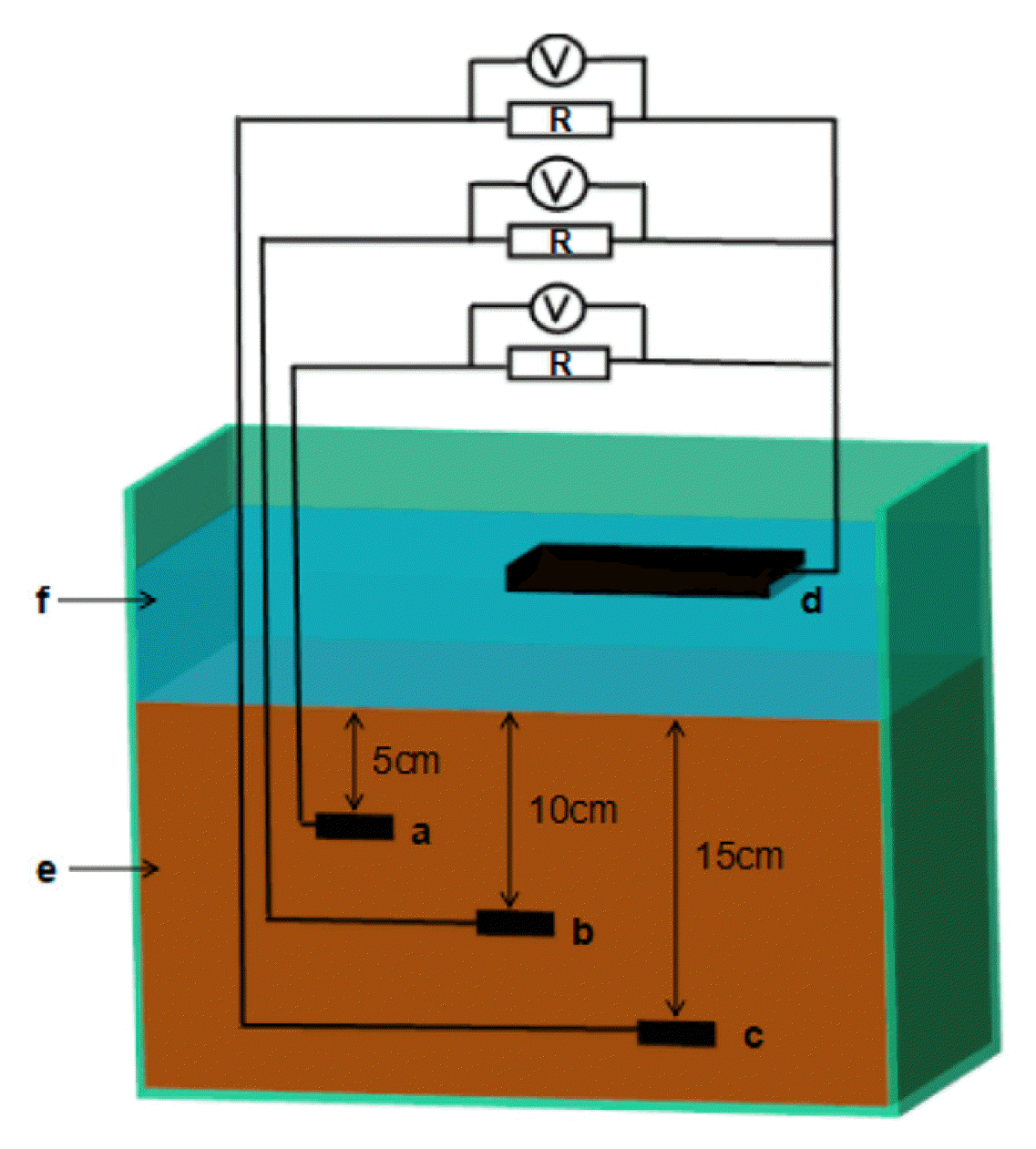
The SMFC reactor was a rectangular container (40 cm×25 cm×60 cm) (Fig. 1). The sediment layer was 20 cm thick, covered by water layer of 7 cm deep. Both anode and cathode were made of graphite felt with a thickness of 0.5 cm. Three anodes with the same size of 4 cm × 6 cm were buried at 5, 10 and 15 cm deep in the sediment, labeled as SMFC-5, SMFC-10 and SMFC-15, respectively. The cathode (20 cm × 12 cm) was placed floating in the water layer and shared by the anodes. The shared cathode was connected to the anodes across an external resistance of 1000 Ω via titanium wire to form SMFC-5, SMFC-10 and SMFC-15. Voltage across the external resistances was recorded at 5 min intervals using a digital data acquisition system (Keithley Instruments 2700, USA).
2.3. Microbial Analysis
After 18 d of operation, the output voltage became stable and implied that the anodic biofilm were matured and ready for bacterial community analysis. Biofilm samples were collected from the anodes surface after the anodes were rinsed with ultra-pure sterile. Bacterial DNA samples were obtaine using a powersoil DNA isolation kit (MO Bio Laboratories, USA) and the collected DNA samples were further purified with a kit (BioTeke, China). The universal primers of 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 533R (5′-TTACCGCGGCTGCTGGCAC-3′) were adopted for the amplification of the V3 region of the bacterial 16S rRNA gene. The PCR amplification procedure followed a protocol in our previous report [24]. The resulted amplicons were then quantitated with QuantiFluor™-ST for further high-throughput sequencing.
Low-quality sequences were removed with QIIME software. The qualified sequences were clustered into operational taxonomic units (OTUs) at 0.03 sequence distance using MOTHUR. Based on the clusters, the number of OTUs, Chao1 index of the rarefaction curves, and phylogenetic richness index for diversity were calculated to determine the diversity and richness of each sample.
3. Results and Discussion
3.1. Effect of Burial Depth of Anode On Voltage Output
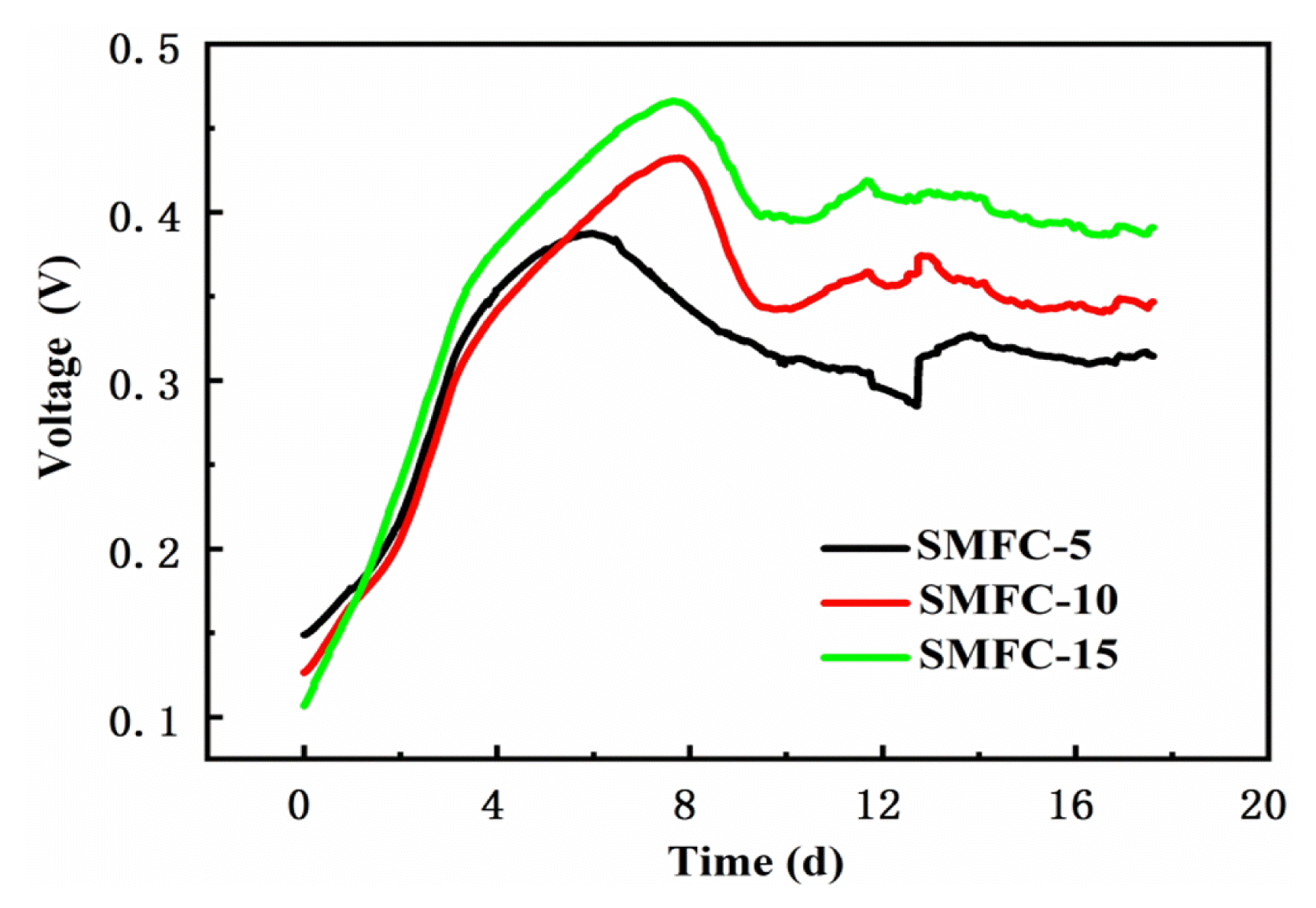
Time-course variations of the voltage in SMFC-5, SMFC-10 and SMFC-15 with an external resistance of 1000 Ω were shown in Fig. 2. It was shown that the voltage output increased quickly in the initial 7 d, and exhibited a peak voltage of 385, 432, 465 mV for SMFC-5, 10, and 15, respectively. Subsequently the voltage decreased and reached a steady state, with an average voltage of 318, 345 and 394 mV for SMFC-5, SMFC-10 and SMFC-15, respectively. The stable voltage output indicated that exoelectronges were successfully enriched on the anode surface [25, 26]. The effect of anode depth on the voltage output was in line with a prior research [1]. The performance of SMFC-15 was better than SMFC-5 and SMFC-10, which implied that the physical and chemical characteristics of deeper sediment favored the metabolism of anodophiles or electrophiles [14, 15].
3.2. Richness and Diversity of Phylotypes
Overall pyrosequencing information and alpha diversity of the bacterial communities are summarized in Table 1. The number of the OTUs obtained was 1438, 1275, 1005 and 2134 for SMFC-5, SMFC-10, SMFC-15 and the original sediment, respectively. The result indicated that SMFC operation and increased burial depth of anode could decrease the microbial diversity. In addition, the maximum number of OTUs were determined as 6271 (SMFC-5), 5008 (SMFC-10) and 3457 (SMFC-15), suggesting that the samples located at deeper depth presented lower richness. The rarefaction curves showed increasing tendency even as the sequencing reads was larger than 3500, which implied that novel microbial phylotypes existed in the bacteria samples (Fig. 3).
3.3. Bacterial Community Structures
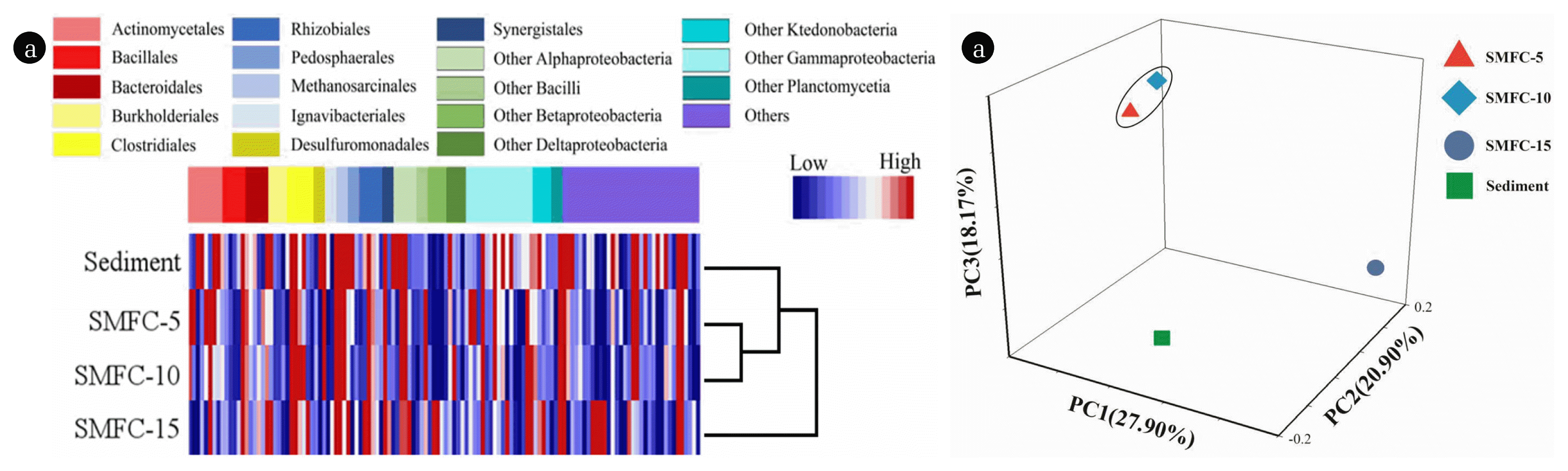
Hierarchical cluster analysis was conducted to reveal the relationship between the bacterial community structures (Fig. 4(a)). It was demonstrated that SMFC-5 and SMFC-10 were clustered together and well separated from SMFC-15 and the sediment. The results were further confirmed by the weighted principal coordinated analysis, for which the components of 1, 2 and 3 accounted for 27.90%, 20.90%, and 18.17% of total community variation, respectively (Fig. 4b). The results suggested that burial depth of anode was an important factor to distinct the bacterial community structure of anodic biofilm in SMFCs.
3.4. Phylotype Distribution
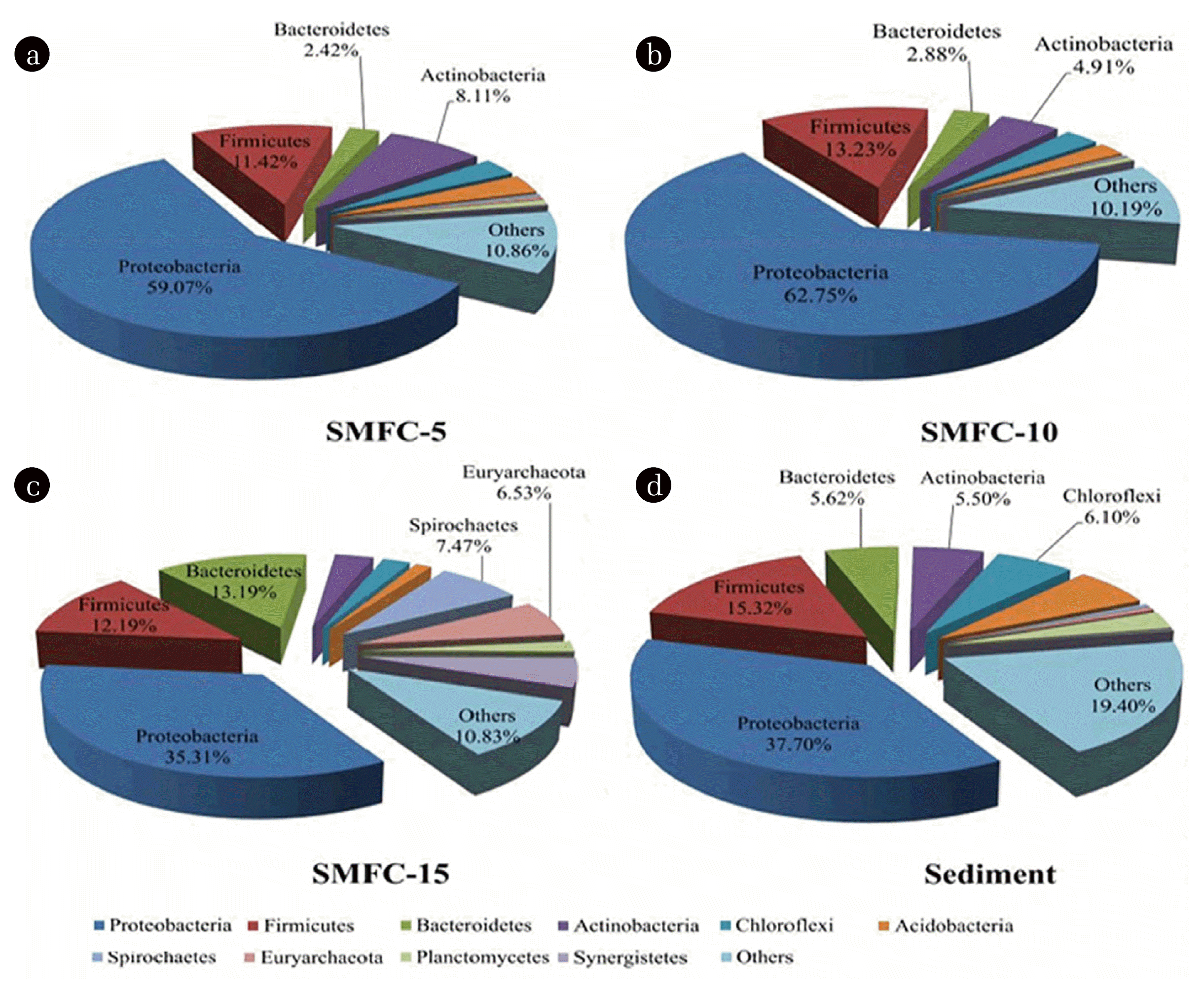
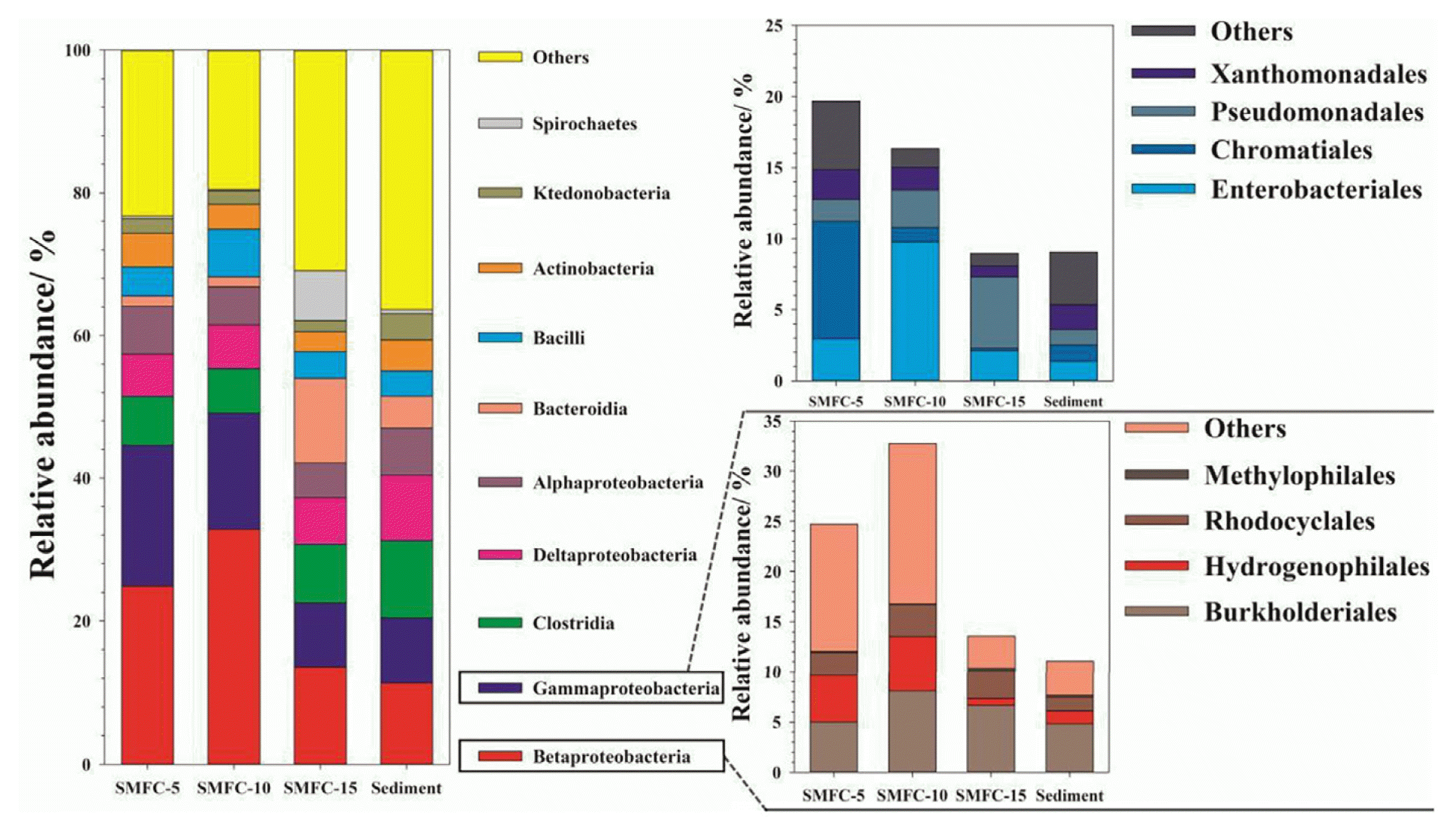
To compare the phylotype distribution of the bacteria samples, the sequences were identified at the levels of phylum, class and order. As shown in Fig. 5, eight dominant bacterial phyla were revealed with a relative abundance > 2%. Proteobacteria has dominated all the biofilms, followed by Bacteroidetes and Firmicutes. They accounted for 72.91%, 78.86%, 60.69% and 58.64% in SMFC-5, SMFC-10, SMFC-15 and sediment, respectively. Bacteroidetes accounted for 13.19% in SMFC-15, higher than 2.42% in SMFC-5, 2.88% in SMFC-10, and 5.62% in original sediment respectively. It was revealed that some members of Bacteroidetes demonstrate the ability of electron transfer to extracellular electron acceptors such as solid anode. The results implied that deeper depth might be beneficial for the accumulation of Bacteroidetes, which enhanced the performance of SMFCs. Chloroflexi is a common filamentous bacteria in wastewater treatment processes, which were abundant in the original sediment (6.10%).
The identification of the bacterial communities at class level in SMFC-5, SMFC-10, SMFC-15 and sediment was illustrated in Fig. 6. Pyrosequencing detected that the majority of sequences belonged to 10 classes. Betaproteobacteria, out of Proteobacteria was determined as the most abundant class. Gammaproteobacteria, another member of Proteobacteria, was revealed as the sub-abundant class in the anodic biofilm, while Clostridia as typical cellulose degradation in a mesophilic environment was the secondly abundant in original sediment (10.78%). Many bacteria out of Gammaproteobacteria, like Shewanella, Citrobacter, Aeromonas, Klebsiella and Pseudomonas, were widely known dominance in MFCs because of their ability to transfer electrons to anodes.
3.5. Characterization of Dominant OTUs
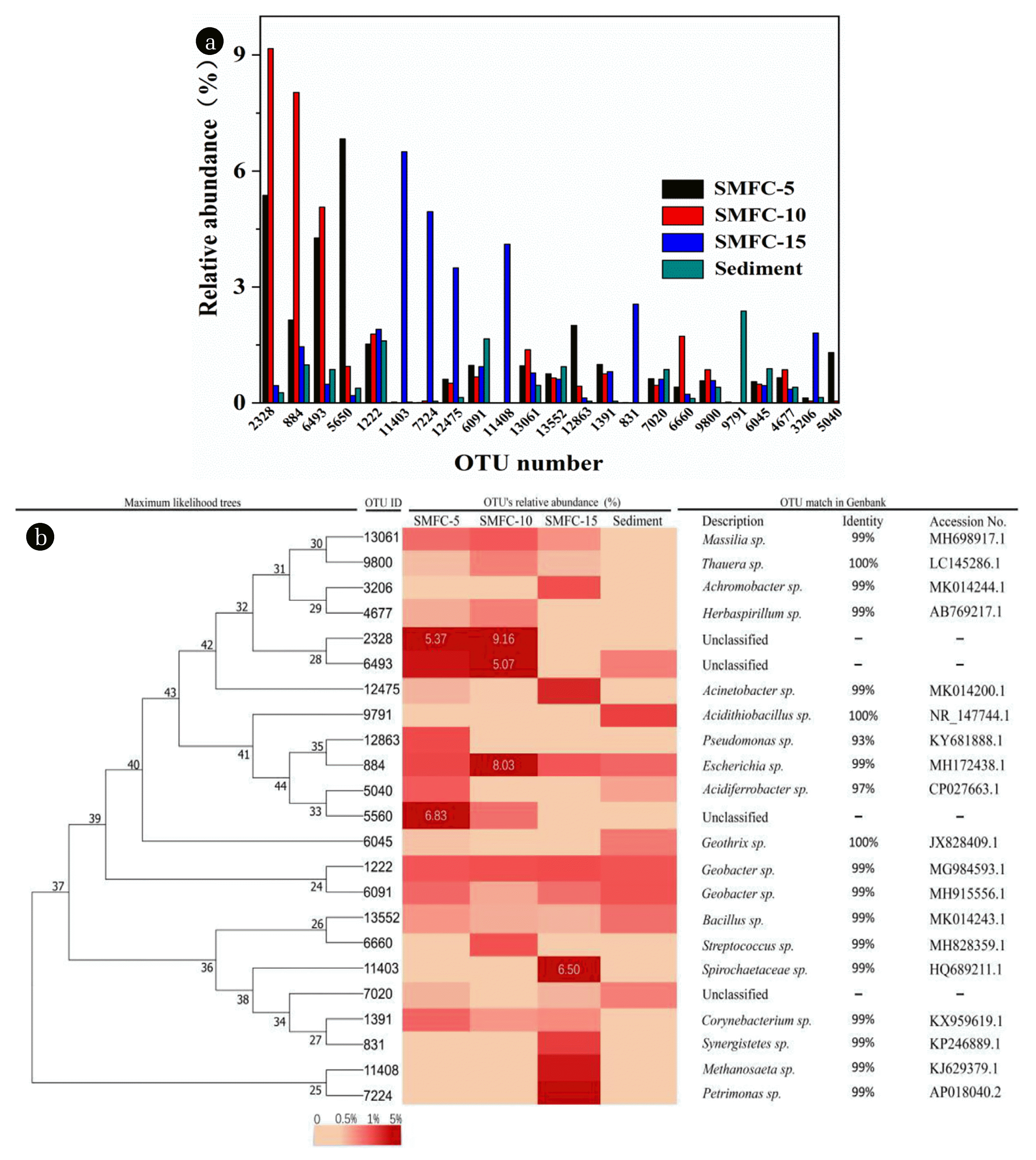
16S rRNA gene libraries were constructed based on the pyrosequencing results. As shown in Table 1, 5861 OTUs were classified with a 3% distance. Out of the 5861 OTUs, only 115 OTUs were shared by the 4 samples. Moreover, 23 OTUs with a relative abundance larger than 0.5% were determined as dominant OTUs and accounted for 13.37–33.94% of the total sequences (Fig. 7(a)). Out of the 23 dominant OTUs, there were 6, 3 and 6 OTUs belonged to the classes of Betaproteobacteria, Deltaproteobacteria, and Gammaproteobacteria, respectively, while only 1 dominant OTU belonged to Bacteroidia, Bacilli, Clostridia, Spirochaetes, and Synergistia. Among the 23 dominant OTUs, three OTUs (2328, 884 and 6493) had a relative abundance larger than 5% in SMFC-10. The results demonstrated that only few species were dominant in the biofilm samples. The relative identifications of the dominant OTUs according to the results of BLAST were illustrated in Fig. 7(b). There were 19 GenBank matches belonging to cultured bacteria, and the other 4 OTUs were unclassified.
The OTUs had different relative abundance in the four microbial communites. The OTU-11403, as the most abundant one in SMFCS-15 (6.50%), rarely appeared in SMFCS-5 (0.026%) and sediment (0.024%), while not found in SMFCS-10. The results indicated that OTU-11403 had higher adaptability in the deeper sediment. The OTU-11403 was phylogenetically related (99% identity) to Spirochaetaceae sp. A previous study demonstrated that Spirochaetaceae had appeared in anodic biofilm of an MFC using dairy manure as substrate [27]. The results indicated that burial depth was an influencing factor to shape the microbial community structure.
Electrochemically active bacteria were an indispensable portion of the SMFC, OTU-1222 and 609 showed similarities of 99% to Geobacter sp. Geobacter sulfur reduces strain is one of the most widely reported electrochemically active bacteria and is known as the model strain for studying extracellular electron transfer mechanism [28, 29]. Besides OTU-1222 and 609, OTUs 12863 was 99% identical to Pseudomonas sp which was also reported as an electroactive bacterium [30, 31].
4. Conclusions
The results demonstrated that the anode installed deeper, produced a higher voltage. The indexes of phylotype richness and phylogenetic diversity of bacteria in biofilm attached to anode were negatively correlated to the burial depth of the anode. Betaproteobacteria and Gammaproteobacteria are more abundant in deeper depth (10 and 15cm). The electroactive microbes of Spirochaetaceae sp. had the largest abundance in the biofilm attached to anode embedded at 15 cm depth. These results further clarified the relationship between current generation and microbial community structure related to location depth of anode in SMFCs, and thus providing valuable information to improve the performance of SMFCs from the perspective of microbial community structure.