1. Introduction
Radon (222Rn), a radioactive inert gas with a half-life of 3.82 d, is generated from alpha decay of radium (226Ra), which is an intermediate of decay of uranium (238U) and thorium (232Th) in the soils and rocks [1, 2]. Radon gas is released into the atmosphere from ground through its diffusion in the pore spaces in soils [3] and it significantly contributes to elevate indoor radon gas concentration through the some mechanisms including convection via cracks and openings, diffusion from soil via the pore space of building materials and emanation from building materials [4]. Thus, soil radon gas can be an effective predictor prone to determine indoor radon concentration. The strong positive correlation between soil radon potential and the percentage of homes above 200 Bq/m3 was established from the survey in Canadian cites [5]. Moreover, the indoor radon concentration in dwellings is governed by ventilation habits, meteorological parameters, individual building characteristics [6], and by the geological and physical conditions of the soil (e.g., soil porosity, tortuosity, water content, particle size and permeability) [7].
In general, radon isotopes decay with emission of alpha particle into charged progenies such as polonium (218Po and 214Po), tending to easily form the molecular clusters, or attach to aerosols or dust particles in the air. Since after, they are inhaled and penetrated deep into the airways of lung and damages DNA of respiratory tissues [8], consequently leading to the respiratory functional changes and even the occurrence of lung cancer to people [9]. In Ireland, the number of patients diagnosed with lung cancer was 3.1 times higher in areas where 10–20% of households were above 200 Bq/m3 of radon concentration, than in which 1% of households were [10].
In the soil environment, the indigenous microbes are inevitable from the ionizing radiations originated from the variety of natural radionuclides such as uranium (238U) and americium (241Am). The ionizing-radiation induce the detrimental effect on the nuclear materials cell DNA [11] through direct (ejection of electron from DNA) or indirect pathways (by reactive oxygen species produced from water radiolysis) [12, 13]. In particular, the gamma radiation has lower ionization ability and higher penetrability comparing to alpha and beta radiation and it can induce a mutagenic effect in microbes, even though the probability of gamma radiation was very low [14]. Bolsunobsky et al. demonstrated that gamma-radiation induced the genetic mutation and simultaneously impaired their growth and survival rate of microbial strains, Salmonella typhimurium and Escherichia coli [13]. Min et al. showed that DNA damage in bacterial cells was positively correlated with the dose of gamma radiation [15]. On the contrast, a few scientific researches proposed that the low-dose of radiation gave the stimulatory effect, increasing function and resistance of the cellular organism [16, 17]. However, to date, this phenomenon has been controversy [18].
Herein, this study aimed to estimate the microbial metabolic activity under the exposure of the varying radon concentrations (i.e., 185, 1,400 and 14,000 Bq/m3) in the laboratory radon chamber equipped with the radon reservoir. Prior to this, the indigenous soil microbes were collected from the different depth intervals, i.e., 0, 15, and 30 cm from radon-contaminated site along with immediately surrounded sites located in the G county, Korea, and then we compared the physicochemical properties of soil and microbial density in focusing on the radon-contaminated soils. Concurrently, their metabolic activity was determined after the radon exposure.
2. Materials and Methods
2.1. Characterization of Physicochemical Properties of Radon-Contaminated Soil
Soil samples were collected in the four sampling sites including radon-contaminated site (R) and three periphery sites (as of controls, C1, C2, and C3) where they are separated by equal distance of 2 m, located in G county, Korea (Fig. 1 and Table 1). At the different depth intervals of 0, 15 and 30 cm below soil surface in each sampling site, samples were taken using a shovel sterilized with 70% (v/v) ethanol solution and then put into a 1 L of amber jar without headspace. Samples were stored on ice and transferred immediately to a laboratory. The radon concentration was measured by Fast Radon Monitor (FRD 400, Radon FT Lab, Korea) in the tightly-sealed container (34 × 24.5 × 17.5 cm, Easy Phillim). Additionally, pH and electrical conductivity of soil samples were monitored by portable meters (Orion star A214 and Orion star A222, Thermo Scientific, Korea) and moisture content, organic matter, and grain-size were also analyzed according to each analytical protocol.
2.2. Estimation of Microbial Activity of Soil Microbes
The microbial activity of soil microbes was estimated by two biological quantification methods, direct colony enumeration and dehydrogenase activity assay. First, for the direct colony enumeration, five grams of soil sample were suspended in 50 mL of sterilized double distilled water and mixed for 10 min at 200 rpm using a separator funnel shaker (MMV-1000W, EYELA, Japan) followed by standing for 30 min. The supernatants were serially 10-fold diluted, and then 20 μL of them were inoculated onto the LB agar plate (composed of Tryptone 10 g/L; Yeast extract 5 g/L; NaCl 10 g/L; and Agarose 15 g/L). After cultivating them at 30°C for 24 h in the static incubator (VS-1203P 1N, Vision Science Co., Ltd., Korea), bacterial colonies were enumerated as of colony forming unit (CFU). Simultaneously, for the estimating the dehydrogenase activity, 1 mL of supernatant was gently-mixed with 10 mL of LB broth into a sterile conical centrifuge tube (FalconTM, USA), and then incubated for 24 h at 30°C. After that, 500 μL of the bacterial culture was put in a 2 mL tube and centrifuged at 10,000×g for 10 min (Centrifuge 5415D, Eppendorf, Korea) to remove remaining nutrient medium, and the cell pellet was suspended with sterile double distilled water. Followed by, it was mixed with 20 μL of 2% (w/v) 2,3,5-triphenyltetrazolium chloride solution sterilized by mixed cellulose acetate syringe filter (0.45 μm, 25AS045AS, Advantec, Japan) and 100 μL of 1% (w/v) glucose (Sigma-Aldrich, USA), and subsequently incubated at 30°C for 6 h (VS-1203P 1N, Vision science, Korea). Finally, 1 mL of chilled methanol was added to extract tetrazolium formazan produced by microbial reduction of 2,3,5-triphenyltetrazolium chloride, after which the absorbance of the extract was determined at 490 nm through the UV-Vis Spectrometer (UV-3300, Humas, Korea).
2.3. Identification of Microbial Strains Based on 16S rDNA Analysis
Microbial colonies cultivating on LB agar plates in the procedure of direct colony enumeration were separately transferred into LB broth and incubated for 24 h at 30°C (VS-1203P 1N, Vision Science Co., Ltd., Korea). The total genomic DNA was extracted by using Fast DNATM SPIN Kit for Soil (MP Biomedical, USA). Polymerase chain reaction was performed using the universal primer set of 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′) and 1492R (5′-TAC GGY TAC CTT GTT ACT T-3′). Amplified PCR products were obtained through the gel electrophoresis (Mupid-α, TAKARA, Japan) using 1% (w/v) agarose in 0.5X TAE buffer (2.424 g Tris base, 0.571 mL acetic acid and 5 mM EDTA in 1 L distilled water), and then they were purified by Dyne Bio Power Gel Extraction Kit (Dyne Bio Inc., Korea). 16S rDNA gene sequence analysis was conducted using an ABI 3730XL DNA Analyzer (Thermo Fisher Scientific, USA). Nucleotide sequences of alignments were performed using Gene bank database on NCBI (National Center for Biotechnology Information: http://www.NCBI.nlm.nih.gov). Microorganism species were acceptably identified based on 95% sequence homology matched to the database.
2.4. Estimation of Microbial Metabolic Activity after Radon Exposure
Soil microbes enriched in LB broth were put into the laboratory radon chamber (1 × 1 × 1 m, acrylic materials) equipped with radon reservoir (RN-1025, Pylon, Canada) and radon measuring instrument (FRD 1600, Radon FT Lab, Korea) and they were exposed to radon radiation with varying radon concentrations (i.e., 185, 1,400, and 14,000 Bq/m3) and exposure time (i.e., 1, 3, and 5 h). Since then, 5 mL of the microbial suspensions were inoculated into the 150 mL of synthetic nutrient solution containing glucose 0.5 g/L; NH4Cl 0.0382 g/L; ZnSO4.7H2O 0.574 g/L; MnSO4.7H2O 0.446 g/L; NaMoO4.2H2O 0.096 g/L; CoCl2.6H2O 0.096 g/L; FeCl3.6H2O 5.0 g/L; AlCl3.6H2O 0.672 g/L; and KI 0.166 g/L. They were subsequently incubated in a shaking incubator at 30°C with the agitation of 160 rpm. Culture growth was monitored with optical density at 600 nm using the UV/Vis spectrometer (UV-3300, Humas, Korea) with direct colony enumeration, and its metabolic activity including enzymatic activity and organic uptake rate were determined by dehydrogenase activity assay (previously described in section 2.2) and chemical oxygen demand (COD) measurement, respectively.
3. Results and Discussion
3.1. Radon Concentration and Physicochemical Properties of Soil Samples
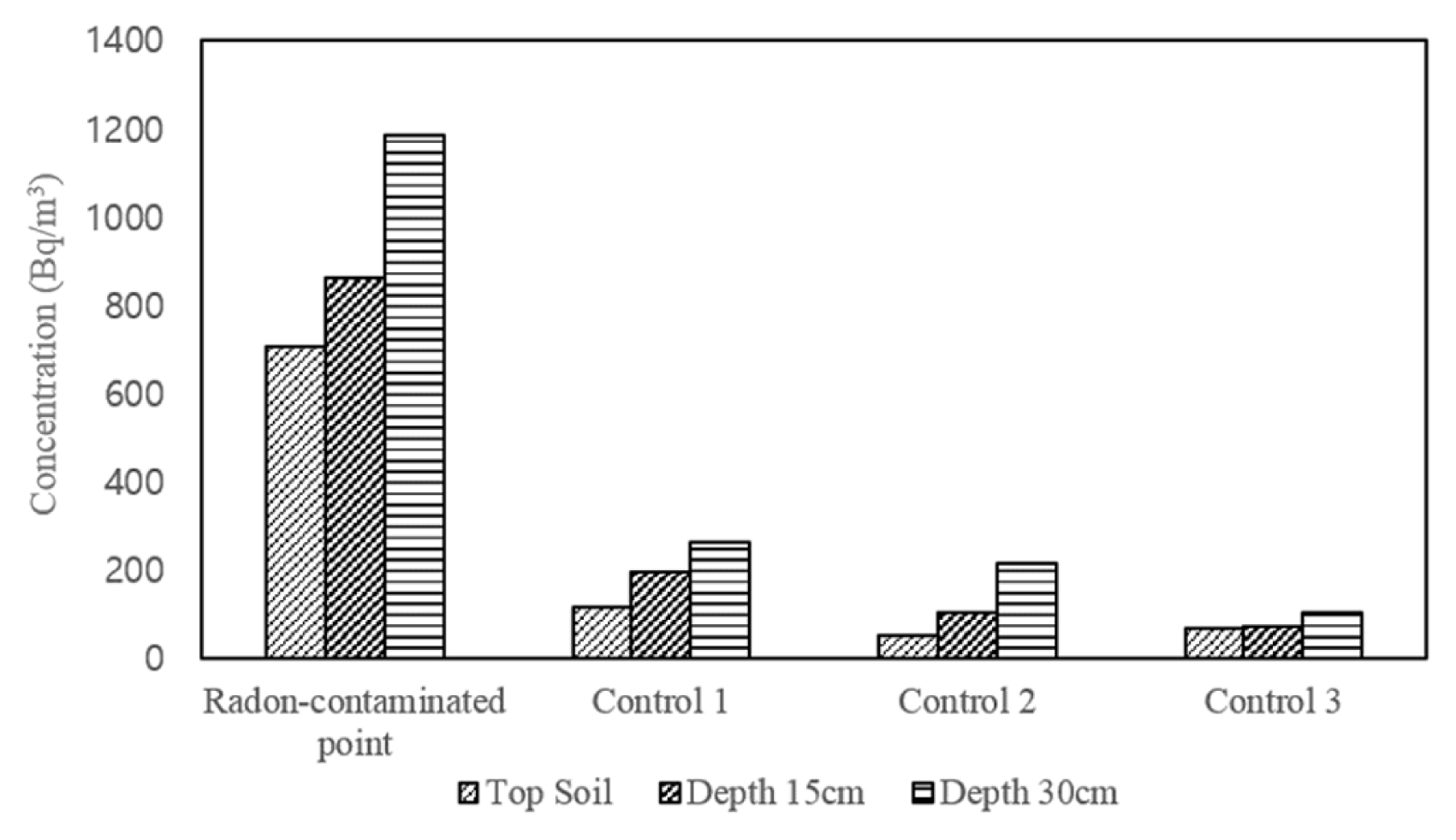
Soil radon concentration tended to decrease with the further distance from the natural radon source (Fig. 2); average radon concentrations on each sampling point were 917 Bq/m3 (R), 192 Bq/m3 (C1), 135 Bq/m3 (C2), and 78 Bq/m3 (C3), respectively. Meanwhile, 707, 860, and 1,185 Bq/m3 was measured at different soil depths of 0, 15, and 30 cm in the radon contaminated soil (R), indicating that the deeper soil has a higher radon radiation intensity than shallower one. The periphery soils (C1, C2, and C3) had a same tendency for deeper soils. The levels of radon concentration were increased at deeper depths because there were more radon sources neighbored in it so that its movement gassing out to atmosphere should be passively limited compared to that of shallower soil depths. It was well known that the atmospheric radon concentration observed near the ground surface was mainly composed of gaseous radon originated from topmost layer of soil and rock surface [7]. Furthermore, referring to Kaur et al. [19], the presence of underlying bed-rocks and the type of soil permeability might ascribe to a difference in radon concentration depending upon sampling sites and soil depths.
Meanwhile, soil radon concentration also depends on the soil physicochemical properties including soil structure, porosity, grain-size, permeability, radium contents, emanation coefficient, and moisture content [20]. In this study, pH, electric conductivity, moisture content, grain-size and organic matter were investigated to compare soil properties between radon-contaminated and periphery sites. First, the pH and electrical conductivity were pH 5.71 ± 0.17 and 0.59 ± 0.06 ds/m in the radon-contaminated soil, respectively and they were comparable with soil samples collected from the C1, C2 and C3 sites, although experimental values in periphery soils were slightly lower. Second, organic matter concentration was also observed no significant difference regardless of either sampling point or soil depths. On the other hand, moisture content tended to decrease with a distance from the natural radon source, similar to radon concentration. It was reported that the moisture content induced the increasing the radon exhalation rates [21], however when the soil pores were saturated by water, the exhalation rate was decreased due to adsorption of radon onto the solid mineral phase or liquid phase [22]. In this study, the moisture content with very broad range from 20% to 60% was relatively higher than other previous studies because the sampling was conducted after rainy spell. Thus, the relationship between the moisture content and radon concentration was difficult to be clearly established. However, the particle size distribution analysis showed that the proportion of sand ranging from 0.02 mm to 2 mm was much higher observed in the radon sources whereas it was declined as far as from them. The sandy soils with more bigger grain size increased soil permeability and consequently contributed to increasing the radon diffusion coefficient [9]. In the meantime, iron species were measured to be higher concentration than those of other metals regardless of sampling sites as can be seen generally on the earth surface.
3.2. Soil Microorganisms at the Radon Source
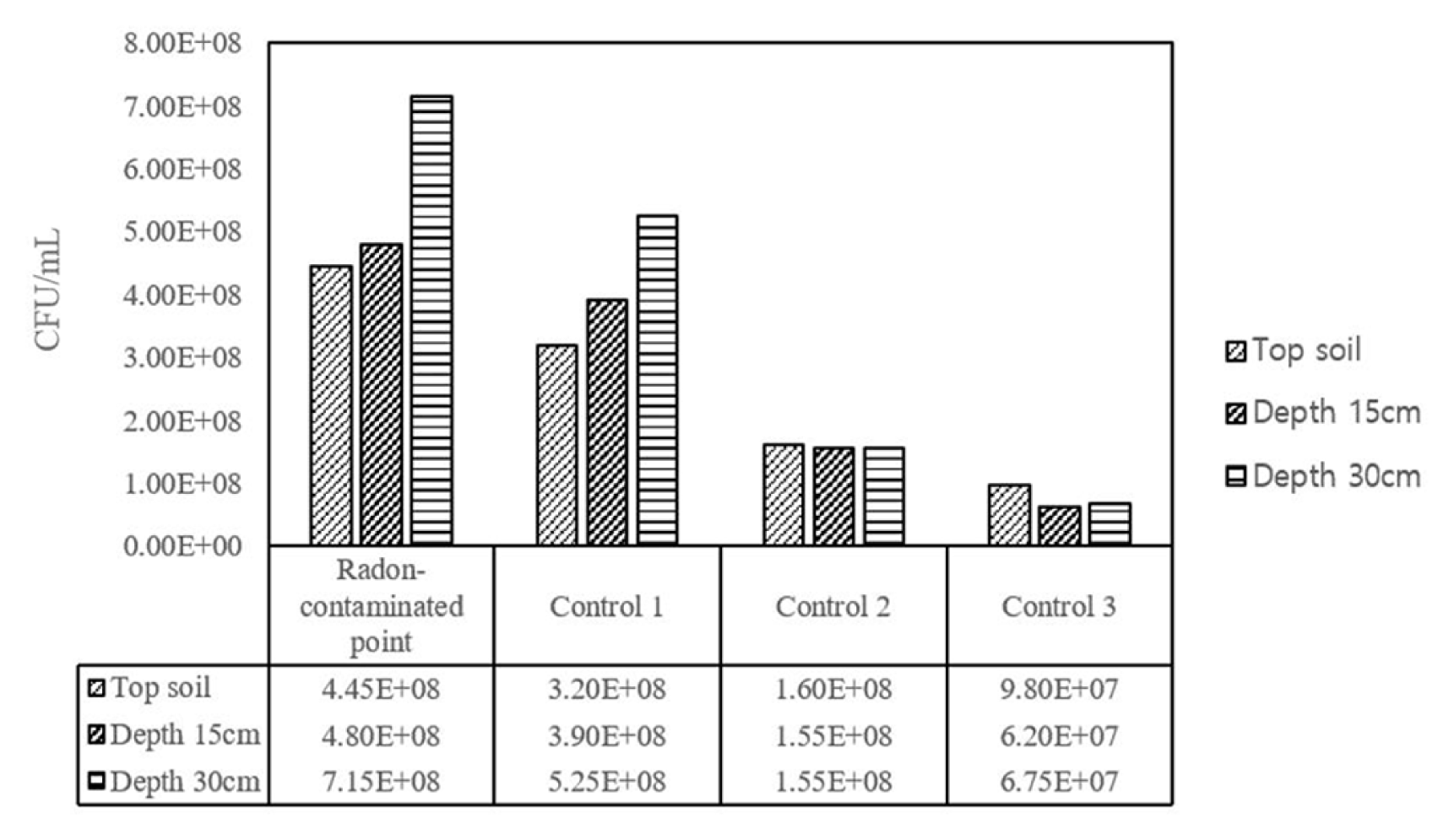
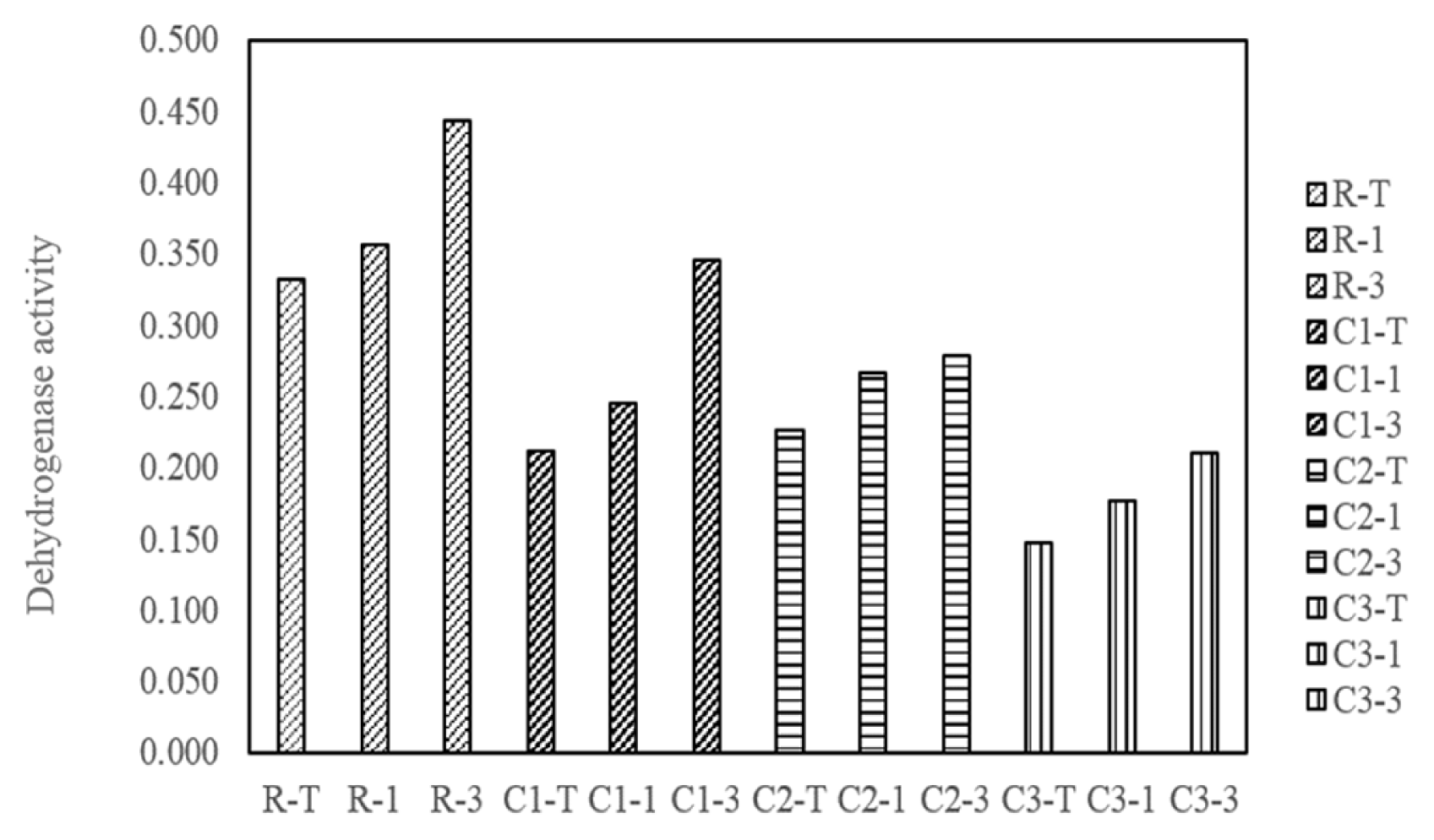
Fig. 3 and 4 showed the microbial population density (as of colony forming in the LB agar plate) and dehydrogenase activity of indigenous soil microorganisms, respectively, observed from both the radon-contaminated and its immediately surrounded soil. In this experiment, as much as the radiation intensity was higher and soil depth was deeper at the study area, the increased number of colonies formed was observed. From this, average of microbial population densities were 5.47 × 108, 4.12 × 108, 1.57 × 108 and 0.76 × 108 CFU/mL for the sampling sites (R, C1, C2 and C3), together with 2.56 × 108, 2.72 × 108, and 3.66 × 108 CFU/mL for the soil depths (0, 15 and 30 cm), respectively. Likewise, dehydrogenase activity was decreased as increasing the distance from the radon natural source, while it was increased as soil depths were increased. This implied that the radon source may have a threshold level that can moreover contribute to proliferating a microbial growth and to exporting diversity at the given site.
Based on 16S rDNA analysis, the indigenous soil microbes were commonly identified by genera of Bacillus, Brevibacillus, Lysinibacillus, and Paenibacillus, regardless of sampling site and soil depths. Each genus included Bacillus cereus, Bacillus subtilis, Bacillus pseudomycoides, Bacillus simplex, Brevibacillus brevis, Lysinibacillus macroides, Paenibacillus pabuli, and Paenibacillus peroriae. Bacillus sp. and Lysinibacillus sp. can grow in the uranium-enriched medium [23], and in particular, Bacillus subtilis adsorbs the uranyl compounds onto their surfaces [24, 25]. Additionally, these microbial strains were known as the potential remediator on the uranium-contaminated soil by sequestrating the uranium [26] or other radionuclides.
3.3. Effect of Microbial Metabolic Activity to Radon Exposure
The variations of microbial diversity and metabolic activity under the radon exposure were investigated on the radon radiation intensities of 185, 1,400, and 14,000 Bq/m3 for 1, 3, and 5 h, respectively. First, the result of microbial diversity of indigenous soil microorganisms based on 16S rDNA analysis indicated that dominant species in the microbial culture exposed with 1,400 Bq/m3 of radon radiation were Bacillus sp. including Bacillus cereus and Bacillus subtilis, regardless of their original habitat environment. Interestingly, Microbacterium sp. and Arthrobacter sp. were more abundantly observed comparing to the not-exposed to radon. These microbial strains, Microbacterium sp. [27] and Arthrobacter sp. [28] has a high resistance to uranium along with the sequestration ability of uranium. These bacterial strains were also observed more predominantly than those of other strains, in the 14,000 Bq/m3 of radon radiation. Mumataz et al. found that the microbes could grow in the medium containing high concentration of uranium in the Ranger Uranium Mine [23].
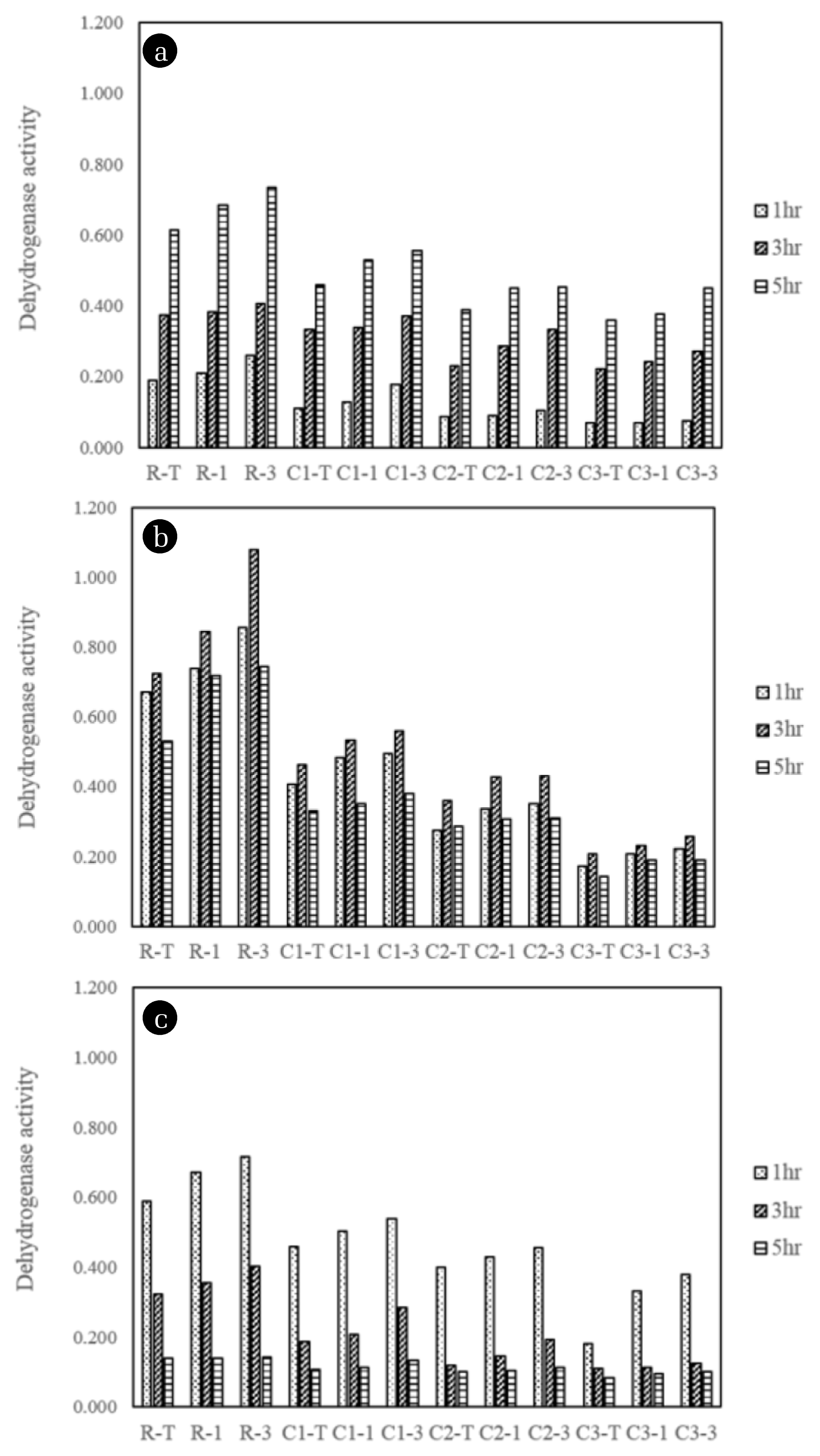
Secondly, the variation of metabolic activity after radon exposure was estimated by monitoring microbial population density (i.e., optical density at 600 nm and colony enumeration), dehydrogenase activity and glucose uptake. Microbial growth of exposed to varying radiation intensities were increased with the incubation time (Fig. S1, S2 and S3) and the maximum microbial density was achieved after 8 to 12 h of incubation. The highest microbial density was found in the experimental sets obtained after 1,400 Bq/m3 of radon radiation employing microbial strains isolated from the radon contaminated soil with its depth of 30 cm (R-3) (Fig. S2(b)). Meanwhile, either lower (185 Bq/m3) or higher radon concentration (14,000 Bq/m3) resulted in the relatively lower microbial density comparing to the radon exposure of 1,400 Bq/m3.
In this context, the result of the dehydrogenase activity (Fig. 5) and direct colony enumeration (Fig. S4, S5, and S6) also showed that the highest number of viable microorganism was detected when the microbial culture was exposed to 1,400 Bq/m3 for 3 h. Simultaneously, Fig. 5 was clearly demonstrated that microorganism isolated from the radon-contaminated soil had a greater metabolic activity than other sampling site where the soil radon concentrations were less than 200 Bq/m3. Meanwhile, the exposure time could be an important factor to determining the microbial activity. At the 185 Bq/m3 of radon concentration, dehydrogenase activity and number of colonies were gradually increased as incubation time was extended from 1 to 5 h. At the 1,400 Bq/m3, the maximum microbial viability was determined in 3 h of radon exposure, and then it was slightly decreased in 5 h. Final radon concentration, 14,000 Bq/m3 had a lower dehydrogenase activity and microbial population density than other experimental condition and steeply decrease of them against increasing exposure time. Hence, it might be implied that the higher radon concentration combined with exposure time above their threshold induced from environmental habitats could exacerbate the microbial activity.
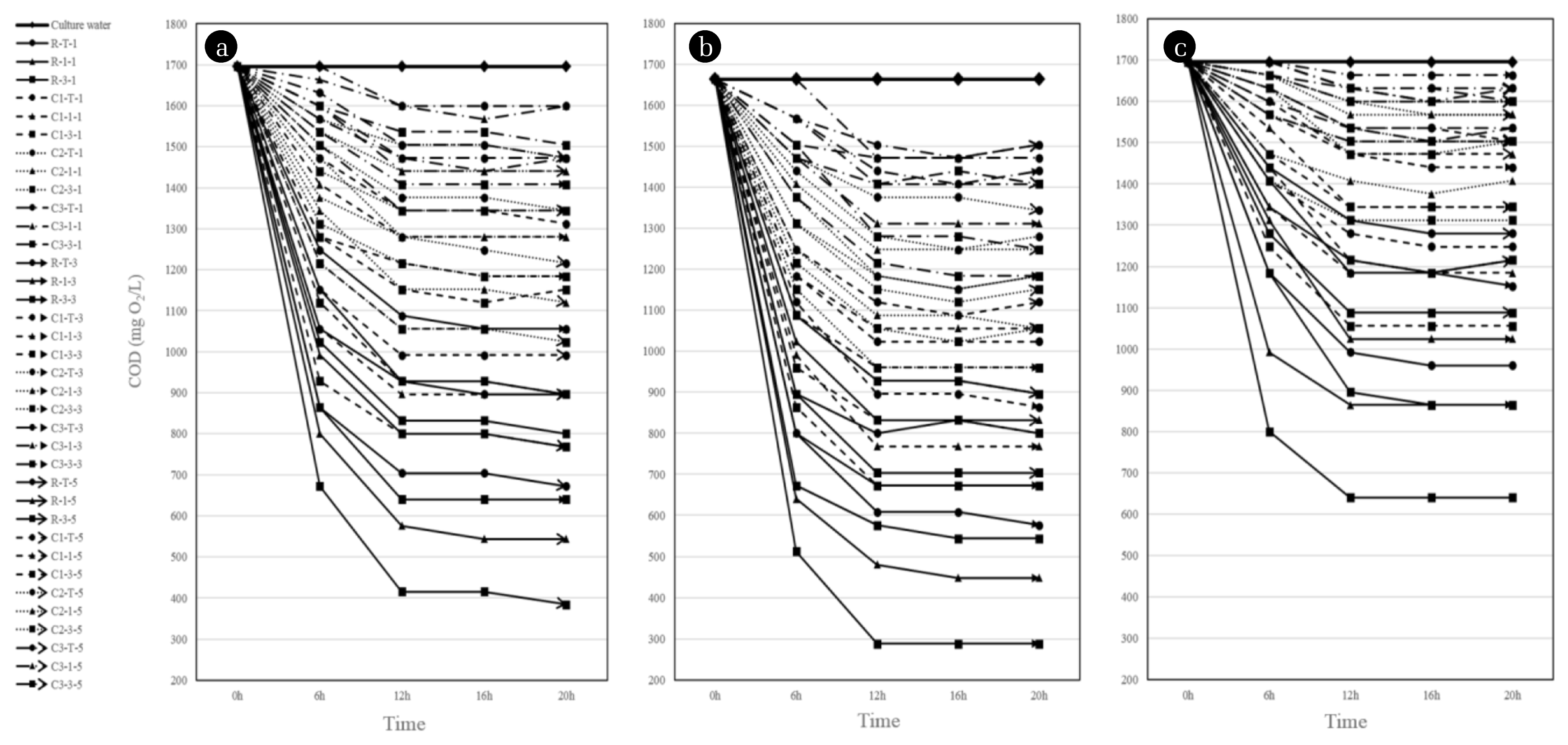
Furthermore, the result of organic uptake rate (Fig. 6) was corresponded with microbial density and dehydrogenase activity. For all microbial culture of experiments, the organic uptake was completed in the 12 h of incubation time, which has no longer significant increase in microbial density, and the maximum organic uptake at this time was slightly different from the radon exposure condition. In particular, the highest organic uptake was observed in the microbial culture isolated from the radon contaminated soil with depth of 30 cm (R-3) after 1 h of radon exposure for each radon radiation intensity. COD value was decreased to 384 mg O2/L (for 185 Bq/m3 of exposure), 288 mg O2/L (1,400 Bq/m3) and 640 mg O2/L (14,000 Bq/m3) from initial concentration of 1,700 mg O2/L, respectively.
Regarding on the whole experimental results of radon exposure, the medium level of radiation (i.e., 1,400 Bq/m3) similar to the microbial habitat of study area could stimulate the microbial growth as well as increase substrate utilization capacity [17, 29]. Whereas, the higher or lower radiation compared to it attributed to the relatively lower metabolic activity of microbes. As aforementioned, high-dose ionizing radiation of radionuclides induce to damage DNA, which consequently bring to the cell mutation or the inhibition of both cell growth and metabolic reaction. It was clearly observed that dehydrogenase activity and organic utilization were more decreased after radon-radiation of 14,000 Bq/m3. While, the low-dose and short time of radiation might be negligible in the viewpoint of the adverse impact of microbial metabolic reaction. It should be present a threshold level for the radiation intensity influencing on microbial resistance. Beyond it, its activity has been seriously impaired, whereas lower than that, it has not been influenced [29]. Furthermore, a few microbial strains are strongly resistant to the radiation of radionuclides. For example, Deinococcus spp. compensates for extensive DNA damage through adaptions such as the genetic redundancy and the enzymatic reaction of specialized proteins that have an ability to protect and repair an impaired DNA following radiation [30, 31]. Some bacterial strains can be viable and metabolically active in the extreme habitation condition akin to the disposal site of radioactive waste, which capable of uranium sequestration [26, 28]. Therefore, it can be inferred that the radiation intensity of radon observed from the study area and tested from the laboratory could give a threshold level for microbes to persistently sustain and moreover proliferlate their viability and population density in a certain range of radiation (i.e., 1,400 Bq/m3).
4. Conclusions
It was the first study to investigate the microbial activity of indigenous soil microorganism against radon exposure. Soil micro-organisms were mainly composed of Bacillus sp., Brevibacillus sp., Lysinibacillus sp. and Paenibacillus sp., and their microbial density and dehydrogenase activity were higher as the closer to the natural radon source. Laboratory radon exposure test with different radon radiation intensities and exposure time showed that microbial activity including microbial population density, dehydrogenase activity and organic uptake was higher in the experimental sets employing the microbial culture isolated from the radon-contaminated soil, same as the result of field study. And the higher level of radon and longer exposure time could diminish the microbial metabolic activity. Consequently, radon radiation could instigate the microbial metabolic activity depending on the radon levels while they were exposed, which could address that the certain extent of threshold concentration present in the ecosystem relevant to microbial diversity and population density.