1. Introduction
Anaerobic digestion (AD) is an organic-waste treatment method that generates energy source methane (CH4). AD commonly operates in mesophilic conditions and requires long degradation time (> 30 d). As a result, organic removal efficiency only reaches 40–70%, and CH4 content reaches only 50–60%. AD is a series of biological reactions performed by various microbial groups [1]. Electron transfer is a vital intermediate in this process; in many AD systems, this function is performed by hydrogen gas (H2) that is generated by bacteria and consumed by methanogens to maintain low H2 partial pressure, but in wet-AD, this process is limited by the slow growth rate of methanogens [2, 3]. DIET may provide an alternative because it does not require energy allocation to multiple enzymatic steps to produce H2. Therefore, DIET increases substrate degradation rate and biogas production [3].
EM is a system that facilitates DIET in AD. EM overcomes thermodynamic constraints by providing external power supply to favor desired reaction [4]. EM operated at 300 mV reached steady-state condition in 50 d and produced CH4 yield 0.34 L/g-CODremoved; whereas control reactor required 100 d to reach steady-state and produced CH4 yield 0.2 L/g-CODremoved [5]. In another study, EM operated at 1,000 mV achieved 2.1 times higher maximum CH4 yields than control reactor [6]. Furthermore, COD removal efficiency of EM operated at 400 mV and 1,000 mV reached 100% within 72 h, whereas conventional AD only reached 56.5% [7]. In summary, previous studies were conducted to investigate the effect of applying voltages on reactors’ performance and the results showed the improvement in CH4 yield and removal efficiency.
Mag (Fe2+(Fe3+)2(O2−)4) supplementation also facilitates DIET in AD. Electron transfer rate facilitated by Mag was reported 106 times higher than control reactors; thereby, enhanced CH4 production by 33% [8]. Under NH3 stress conditions, Mag supplementation increased specific methanogenic activity (RCH4) of reactors fed with acetate, propionate, and butyrate by 58%, 20%, and 15%, respectively [9]. Previous studies were conducted to quantify the effect of Mag supplementation in methanogenesis, and the results showed the enhancement in CH4 production.
Adding Mag to EM system has also been proposed to improve the conductivity around electrode surfaces. Mag facilitates electron transfer from electroactive microbes located far from electrode surface because it acts as electron conduits [10]. The costs of implementing EM systems are being reduced by development of inexpensive electrode materials and efficient system configurations [4]. Also, Mag can be recycled for long-term continuous AD (≥ 317 d) without compensating DIET activity or methanogenesis efficiency [11].
Still, adoption of EM systems coupled with Mag supplementation requires optimization of EM conditions with Mag supplementation under various substrate conditions that the field may encounter, such as the presence of NH3. EM or AD system coupled with Mag supplementation has been investigated under optimal conditions [1, 4, 5, 7, 12]. However, protein degradation produces NH3 as a byproduct, and proteins usually comprise 40–70% of total volatile solid in AD substrates, so [NH3] can build to levels that inhibit the activity of methanogenetic consortia [13, 14]. A certain amount of [NH3] (< 200 mg/L) is beneficial for microbial growth, but high [NH3] may restrict it [13]. NH3 diffuses passively into microbial cells and thereby increases the intracellular pH. To counteract this effect, cells must devote energy to proton balancing by using a K+ pump. Therefore, it increases the maintenance energy needed to survive in high [NH3] and potentially inhibits other enzymatic reactions involved in methanogenesis [15].
There is lack of study regarding the optimum operational condition of EM coupled with Mag supplementation under different NH3 conditions. Therefore, this study was conducted to investigate the effect of VCAT and [Mag] under different NH3 conditions on methanogenesis of acetate. Acetate is an important intermediate that contributed to two-third of produced CH4. Considering the roles of acetate as a pivotal intermediate and methanogenic precursor consolidates its importance in AD [16]. This study was also conducted to investigate how independent variables affect microbial growth. This study is significant to identify conditions that are beneficial for full-scale operation of an EM system that uses Mag supplementation.
2. Material and Methods
2.1. Inoculum Source and Media
The microbial seed was collected from 3.5 L (working volume) continuous lab-scale mesophilic anaerobic reactors operated for two years, treating sludge and foodwaste. Throughout the operation of inoculum system, [TAN] was < 1.5 g/L and 14.9 g-VSS/L. The inoculum source had never been exposed to any bioelectrochemical systems nor supplemented with any semi(conductive) materials. Before inoculation, the seed was starved for seven days at 37°C. Anaerobic basal media was prepared as previously described [17]. The growth medium was supplied with 2 g-COD acetate/L as substrate. Different concentrations of ammonium chloride 2.2, 11.7, and 21.3 g/L and 1.3 g/L sodium bicarbonate were supplied as NH3 nitrogen and bicarbonate sources.
2.2. Construction of EM Reactor and Operational Conditions
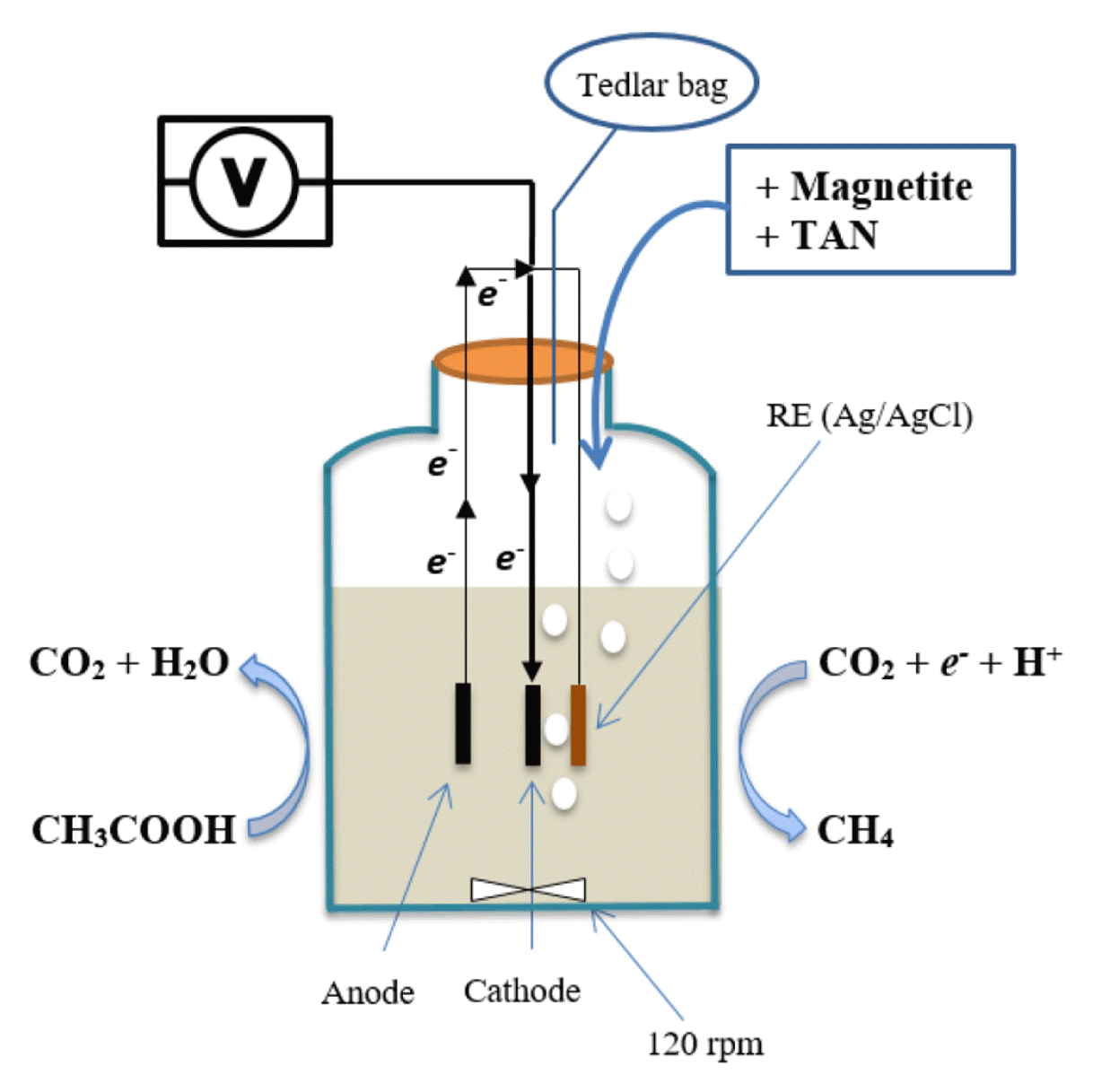
Single-chamber membrane-less systems were constructed for this study and operated in batch mode. The EM reactors (Fig. 1) were composed of 320-mL glass bottles (HwaIn Auto Living Co., China) with 180-mL working volume. Electrodes were treated and prepared as previously described [18]. The anode was 3.5 cm × 2.0 cm (length × width) isomolded graphite plate (Grade GM-10; GraphiteStore.com, Inc., USA). The cathode was stainless steel mesh (type 304, mesh-size 90 × 90; McMaster-Carr, USA) cut in the same dimension as anode. The reference electrode (RE) was Ag/AgCl RE-21B EC-Frontier, Japan (+210 mV vs. standard hydrogen electrode, SHE). Voltages in this manuscript were reported with respect to SHE. Electrochemical monitoring was performed by multichannel Potentiostat/Galvanostat WMPG1000LeK8 (WonATech, Korea).
The reactors were supplied with basal media containing substrate and 1 g-VSS/L inoculum source, then purged with 99.9% nitrogen gas. The medium’s pH was maintained at 7.5±0.1 by adding 2.5-N NaOH or 2-N HCl as required. The temperature was maintained at 37±1°C. The reactors were operated under 120 rpm continuous stirring. Experiments were conducted until biogas production ceased and the substrate completely degraded.
2.3. Experimental Design
A central composite design face-centered (CCF) with full 33 factorial design was used, because the region of operability covers the region of interest. VCAT X1, [Mag] X2, and [TAN] X3 were independent variables applied (Table 1), and their ranges were determined in preliminary experiment (data not shown). To evaluate the models’ error, center points (0, 0, 0) were run in triplicate. Seventeen experimental runs were conducted (Table 2). Statistical analysis of CCF was performed using Minitab 17.1.0, PA-USA. The observed responses were cumulative CH4 production Y1 and acetate degradation rate Y2. Second-order polynomial model, Eq. (1) was used to analyze and fit experimental results obtained from CCF design [19].
where Y is predicted response; β0 is constant term; k represents number of variables; βi, βii and βij represent coefficient of linear, quadratic, and interaction effect; xi and xj are coded values variables i and j.
VCAT was selected considering over potential and electromotive force that favors CH4 production [20, 21]. Acetate as simple substrate was supplied to avoid any side reactions that may occur at −1,000 ≤ VCAT ≤ −600 mV, which can decrease CH4 production [22]. [Mag] added to the systems was selected considering previous studies [12, 23]. [TAN] was selected considering inhibitory and non-inhibitory level: 1.5 g-TAN/L is accepted as non-inhibitory level, thus, used as low concentration; 4 g-TAN/L that reduces methanogenesis by 40–60% was chosen as center point; and 6.5 g-TAN/L that reduces methanogenesis by 80–100% was set as maximum [TAN] [13, 14]. Maximum [TAN] used in this study also adopted from maximum [TAN] observed in ten full-scale anaerobic digesters treating different types of protein-rich substrates [24].
2.4. Real-time Quantitative Polymerase chain Reaction (qPCR) Analysis
Samples for qPCR analysis were taken at the beginning and endpoints of experiment. Total DNAs of the samples were extracted using a fully-automated nucleic acid extractor (Magtration System 12GC, PSS-Co., Japan). 100 μL Tris-HCl buffer (pH 8.0) was added for elution of the extracted DNA. The eluted DNA preserved at –20°C for further analysis. TaqMan probe-primers, as previously reported [25], were used to quantify population of total bacteria and archaea, Methanosaetacea, Methanosarcinacea, Methanomicrobiales, Methanobacteriales, and Methanococcales using qPCR analyzer (LightCycler 480-Roche, Switzerland).
2.5. Physicochemical Analysis
Gas volume was measured using water displacement method. Biogas contents were analyzed by gas chromatograph (GC-HP 6890, Agilent, Palo Alto, CA-USA), equipped with HP-Plot Q packed column and thermal conductivity detector. Helium (He) used as carrier gas with 20 mL/min flow rate. Acetate concentrations were analyzed using gas chromatograph (GC-HP 6890 Plus Agilent, Palo Alto, CA-USA), equipped with Innowax capillary column (Agilent) and flame ionization detector. The carrier gas was He with 10:1 split ratio and flow rate 2.5 mL/min. Standard methods protocols [26] were applied to measure [TAN] using Kjeldahl method. pH was measured using a pH electrode (Cole-Parmer Instrument). All analytical items were analyzed in triplicate, and results represented as mean±standard deviation.
2.6. Statistical Analysis
Acetate degradation rate was estimated using first-order kinetic model:
where [Acetate]t (g-COD/L) is acetate concentration at t observation time; [Acetate]0 (g-COD/L) is initial acetate concentration; k (d−1) is first-order kinetic coefficient for acetate degradation rate; and t (d) is observation time. Principal component analysis (PCA), analysis of similarity (ANOSIM), and Pearson r correlation were performed using statistical language R-4.0.2.
3. Results and Discussion
3.1. Model Accuracy and Validation
CCF design was used to quantify the combine effects of independent variables on methanogenesis of acetate under different NH3 conditions. The following quadratic equations illustrate the mathematical-statistical relationship between factors and responses in coded units.
where Y1 represents cumulative CH4 production, Y2 represents acetate degradation rate, X1 is VCAT, X2 is [Mag], and X3 is [TAN]. The coefficient of determination R2 indicates the accuracy of polynomial equality [27]. Calculated R2 = 0.950 for Y1 and 0.903 for Y2 denote that the models explain 95% of cumulative CH4 production and 90.3% of acetate degradation rate.
Response surface plots (Fig. 2) drawn using Eq. (3) and (4) illustrate the function of cumulative CH4 production and acetate degradation rate when [TAN] was held at center point (4.0 g/L). The maximum value of both responses is located inside the experimental region (Fig. 2a, b). The adequacy of the models was confirmed by ANOVA results (Table 3). Both response models were significant (p < 0.05) and had insignificant lack of fit (p > 0.05) that indicates the models were fitted well to the experimental data and can be used to predict the responses [19].
Response optimizer goals were set at maximum point to determine optimal conditions of independent variables with maximum responses. A validation trial was performed to assess the models by comparing the predicted and actual values at the identified desirable conditions. The difference between predicted and actual results was 1.86% for Y1 and 4.63% for Y2 (Table 4). Up to 5% difference between predicted and actual values is acceptable to confirm the adequacy of response surface models [27].
Optimized VCAT = −737.4 mV for Y1 and −757.6 mV for Y2 are more than enough to drive reactions (Eq. (5) and (6)) on cathode surface, which at least requires −650 mV. More negative VCAT is required to accommodate ohmic losses [28]. The result is aligned with previous study that reported −600 ≤ VCAT ≤ −800 mV improved CH4 production and substrate degradation in AD systems [29].
VCAT significantly affected cumulative CH4 production (Table 3), as shown in previous study [29]. In EM system, cathode acts as electron donor that is essential to control CH4 production, CH4 yield, and reaction kinetics [30]. VCAT also affects the availability of electrons at the cathode and ultimately also affects the expression of genes related to CH4 metabolism. For instance, the expression of methyl-coenzyme reductase (Mcr) reported 50% higher at VCAT = −1,000 mV vs. Ag/AgCl (−800 mV vs. SHE) than at VCAT = −700 mV vs. Ag/AgCl (−500 mV vs. SHE). Mcr catalyzes the final step of CO2 reduction to CH4, so decrease in Mcr would decrease CH4 production [31]. Therefore, applying appropriate VCAT is essential to favor EM condition. These are the reasons that VCAT was selected as critical parameter in this study.
Optimized [Mag] was 18.2 mM for Y1 and 21.4 mM for Y2. These results confirmed the result of previous study [23] that investigated the effect of [Mag] on cumulative CH4 production and CH4 production rate until 320 mM. The authors stated that 20 mM of Mag enhanced methanogenesis of volatile fatty acids (VFAs), and further increase of [Mag] did not improve the reaction rate.
The optimized condition of [TAN] for both responses was at 1.5 g/L, which is accepted to be non-inhibitory [13]. TAN has no adverse effect on methanogenic consortia when it accumulates up to 1.5 g/L. Above this level, intolerable free NH3 nitrogen concentration ([FAN]) may accumulate in the digester and inhibit methanogenesis [15].
[TAN] significantly affected CH4 production and acetate degradation rate (Table 3). From the equilibrium equation of [TAN] and [FAN] [32], the calculated [FAN] supplied in this study were 59, 157, and 256 mg/L, which correspond to 1.5, 4.0, and 6.5 g/L TAN, respectively. The inhibition effect of TAN is caused by the diffusion of free NH3 (FA) through the membrane cell; thus, cells must increase the energy invested to maintain intercellular pH. Eventually, the expression of several key enzymes that contribute to substrate degradation and product formation will be disrupted [15, 33]. For methanogens, FAN substantially suppresses the expression of coenzyme F420, which is a key coenzyme for CH4 production, up to 97% after exposure with 254 mg-FAN/L for 50 d [34]. These results explained the significant effect of [TAN] on both responses and the low cumulative CH4 production and acetate degradation rate when the reactors were supplied with 6.5 g-TAN/L.
3.2. Interaction between Independent Variables
The signs of the coefficients indicate promotion or inhibition of each independent variable on the response functions [27]. Eq. (3) and (4) both indicate that Y1 and Y2 are negatively correlated with VCAT and [TAN] but positively correlated with [Mag]. These indicate that the more negative VCAT and the higher [TAN] inhibit cumulative CH4 production and acetate degradation rate, whereas the increase in [Mag] promotes both responses.
VCAT and [TAN] showed significant interaction (p < 0.05) on Y1 and Y2. VCAT supplied to the system provides electromotive force for NH3 removal: ammonium ions that react with hydroxides in the system are converted to NH3, which then escape into the headspace of the reactor. This is the alleviative effect of EM in NH3 inhibition conditions during AD [35] that also explained the interaction between VCAT and [TAN] on both responses.
In contrast, [Mag] did not interact significantly with either VCAT or [TAN] (p > 0.05). This result indicates that [Mag] had less effect than other independent variables. Mag has been reported responsible for reactor stability operated over 600 d [36]. Therefore, Mag supplementation showed insignificant effect on two responses for linear, quadratic, and interaction terms due to short-term operation. This also indicates that VCAT alone is sufficient to promote reaction rate through DIET during short-term operation.
Regardless of the insignificant effect of [Mag] in linear and interaction terms, Mag alleviates NH3 stress in AD indirectly. Microbial cells require more energy for cell maintenance to survive in high [NH3] that potentially inhibits other enzymatic reactions involved in methanogenesis [15]. A high electron-transfer efficiency mediated by Mag alleviates the problem of energy deficiency caused by NH3 inhibition [9].
3.3. Effect of VCAT, [Mag], and [TAN] on Microbial Populations
PCA was performed on total bacterial and archaeal populations to investigate how microbial populations were affected by independent variables employed in this study (Fig. 3). The bacterial and archaeal populations were positively correlated (r = 0.63, p < 0.05), and first two principal components (PC-1 and PC-2) correspond to 83.1% of the total variation between microbial populations. The microbial populations clustered based on VCAT regardless of [Mag] and [TAN] supplied to the systems. To confirm the significant difference between those clusters, ANOSIM was performed with Euclidean distance and 9,999 permutations. The ANOSIM R-value of the test was 0.611 (p < 0.01), indicates that microbial growth varied significantly in response to VCAT.
Violin-boxplots (Fig. S1) visualize VCAT influence on total bacterial and archaeal populations. At the beginning of the experiment, average populations were 1.19 × 109 (±2.24 × 108) copies/mL for total bacteria and 1.29 × 107 (±1.61 × 106) copies/mL for total archaea.
The effect of VCAT on microbial populations varied according to its magnitude. At the end of experiment, VCAT = −600 mV contributed to significant increase (p < 0.01) of microbial populations with average total bacteria and archaea were 1.32 × 109 (±4.16 × 108) copies/mL and 1.47 × 107 (±1.07 × 106) copies/mL, respectively. The supplies of different [Mag] and [TAN] led to a wide distribution of bacterial populations. In contrast, reactors that were subjected to VCAT = −800 and −1,000 mV, the different [Mag] and [TAN] supplied to the system had comparatively little effect on microbial concentration, which is indicated by its narrow distribution. VCAT = −800 mV significantly increased (p < 0.01) total microbial concentration to almost double at the endpoint of experiment, with 2.10 × 109 (±7.29 × 107) copies/mL of bacterial and 2.04 × 107 (±1.71 × 106) copies/mL of archaeal concentrations. Whereas VCAT = −1,000 mV caused significant decrease (p < 0.01) of microbial population with (4.28±1.68) × 107 copies/mL and 1.38 × 106 (±5.79 × 105) copies/mL of bacterial and archaeal concentrations, respectively.
Appropriate VCAT may increase the abundance of several functional features, such as biogenesis of cell wall/membrane/envelope, coenzyme transport and metabolism, and citrate cycle metabolic pathway [35]. These functional features are responsible for the improvement of extracellular electron transfer that is involved in methanogenesis. This relationship explains the increase of microbial concentrations in reactors that had been subjected to VCAT = −800 mV, which led to high cumulative CH4 production and high acetate degradation rate (Fig. 2).
In this study, electrochemical NH3 removal was only observed in reactors to which VCAT = −1,000 mV ([TAN] decreased 73.7±5.5%) or −800 mV ([TAN] decreased 6.2±1.1%) had been applied. This observation also aligns with previous study [27] that reported up to 70.6% decreased [TAN] in their EM system. Increasing negativity of VCAT could trigger rupture of cell wall and cause irreversible damage to cell membranes [37]; this response explains the decrease of microbial concentrations at VCAT = −1,000 mV (Fig. S1), regardless of the alleviative effect of strong VCAT in reducing [TAN]. Furthermore, decreasing microbial concentrations caused low cumulative CH4 production and acetate degradation rate (Fig. 2).
The final product of acetate degradation in this study was CH4. The acetate-removal efficiency of all reactors reached 100% after different elapsed times because it affected by independent variables supplied to the system. Regardless of [Mag], cumulative CH4 production at VCAT = −600 mV and −800 mV with [TAN] = 1.5 g/L and 4.0 g/L ranged from 78.8–118.0% of theoretical value. Cumulative CH4 production > 100% of theoretical value in EM system has been reported previously [7]. Considering some fraction of substrate is utilized by microorganisms for cell synthesis and maintenance [38], the result indicated that acetate was mainly utilized for CH4 production. In contrast, CH4 production at VCAT = −1,000 mV with 6.5 g-TAN/L ranged from 6.4–40.3% of the theoretical value. The acetate removal efficiency also reached 100% in this system, despite the low CH4 production. The results indicate that the acetate was mainly used for cell maintenance to survive in NH3 stress conditions.
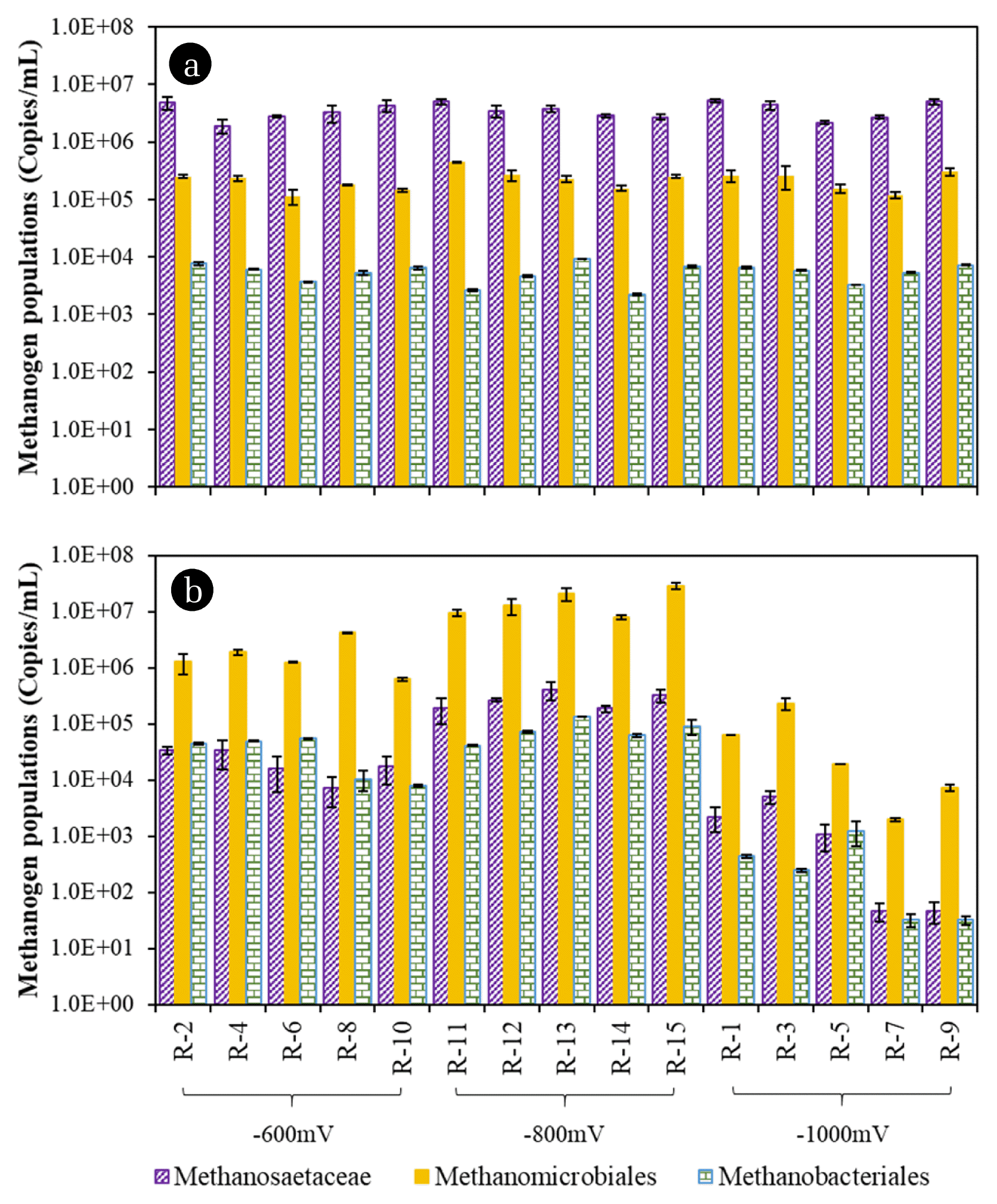
Methanogen populations were also quantified to identify the dominant methanogens (Fig. 4). Of three orders (Methanococcales, Methanobacteriales, and Methanomicrobiales) and two families (Methanosarcinacea and Methanosaetacea) that commonly occur in anaerobic digesters, only Methanosaetacea, Methanomicrobiales and Methanobacteriales were detected. As shown in Fig. 4, VCAT shifted methanogenesis pathway from acetoclastic to hydrogenotrophic. It was indicated by the decrease of Methanosaetaceae concentration and increase of Methanomicrobiales and Methanobacteriales at end of experiment.
Although microbial populations were mainly affected by VCAT in this study, [Mag] and [TAN] also affected them to some degree. Mag affects methanogens growth and its activities [12, 39–41]. Population of hydrogenotrophic methanogens (HMs), especially Methanobacteriales increased by > 12-times in 376 d. Mag stimulated the growth of methanogens through syntrophic VFA-oxidizing bacteria via DIET [12]. Furthermore, Mag supplementation enhanced methanogenesis by increasing the expression of coenzyme F420 and coenzyme M by up to 1.8 and 1.7-times, respectively. Relative abundance of Methanolinea, which belongs to Methanobacteriales, increased up to 62.6%. Excitation emission matrix fluorescence spectroscopy also showed significant increase of soluble microbial products (SMPs) in semi-continuous AD supplemented with Mag for 20 d [39]. SMPs are organic compounds released during normal microbial metabolism; they indicate the activity of AD systems [40]. Another study reported the increase of mcrA gene copy numbers up to 4.3-times in batch AD after supplemented with Mag for 29 d [41].
[TAN] from 1.7 to 14.0 g/L has been reported to reduce methanogenesis by 50%. The wide range of [TAN] inhibition influenced by operational conditions (pH, temperature, HRT), substrate conditions, and inoculum acclimatization [13, 14]. TAN affects the growth and activities of methanogens and the growth of syntrophic VFA-oxidizing bacteria, although the inhibition effects are less severe than methanogens. Therefore, AD system suffering from NH3 inhibition will also suffer from VFA accumulation [15].
HMs have better tolerance than acetoclastic methanogens to high NH3 levels [13, 14, 42, 43]. HMs can maintain homeostatic system by activating K+ or Na+/H+ antiporter and K+ uptake systems to regulate turgor pressure and maintain cytoplasmic pH as a response to NH3 protonation. Concurrent translocation of Na+/H+ across the membrane leads to proton gradient generation that is required to synthesize adenosine triphosphate. Therefore, HMs can conserve energy in NH3 stressed conditions [44]. This ability may explain the increase of HMs population in this study.
This study provides valuable information to guide optimization of an EM process coupled with Mag addition in the presence of NH3. Nevertheless, this study was conducted in batch system. Further experiments should consider process optimization of similar independent variables in continuous system that uses complex substrates.
4. Conclusions
The interactions of VCAT and [Mag] on cumulative methane production and acetate degradation rate in EM system at different NH3 conditions were quantified using a response-surface model. The optimal responses that enhanced cumulative CH4 production were achieved at VCAT = −757.6 mV, [Mag] = 18.2 mM, and [TAN] = 1.5 g-TAN/L. The optimal responses that enhanced acetate degradation rate were achieved at VCAT = −737.4 mV, [Mag] = 21.4 mM, and [TAN] = 1.5 g-TAN/L. VCAT applied to the system promoted microbial growth (at VCAT = −800 mV) or electrochemical NH3 removal (at VCAT = −1,000 mV). VCAT application also caused a shift in methanogenesis pathway from methanogenic to hydrogenotrophic.