1. Introduction
Estrogens are essential group of hormones that regulate the growth and development of female sexual characteristics in humans and animals. Estrogens strongly interfere with endocrine and reproductive functions in living organisms including disturbance on immunological system and fertility, reproductive failure due to thinning of eggshells and reduction in egg production [1], feminization and masculinization, and altered sexual development [2]. 17α-ethinylestradiol (EE2), a synthetic estrogen, is generally used as the oral contraceptive pill to control the birth rate and hormone therapy. EE2 prompt to be released to the environment via human excretion which is later discharged into municipal wastewater treatment plants (WWTS) as hotspots for the release of estrogens in the environment [3].
Some estrogens are able to be degraded in the WWTS. The estrogens removal efficiencies in activated sludge process range from 61% to 98% for estrone (E1), 67% and 99% for estradiol (E2) and 85% to 90% for EE2 [4]. Natural estrogens (E1 and E2) are readily metabolized by heterotrophic microorganisms in WWTS, while the biodegradation of more recalcitrant hormone like EE2 is known mainly through cometabolism of ammonia mono-oxygenase enzyme (AMO) by ammonia-oxidizing bacteria (AOB) [5, 6]. Some essential factors like the availability of ammonia (NH3) as the growth substrate of AOB [5] have been investigated for its roles on EE2 cometabolism by AMO. However, microorganism generally produce enzyme and other macromolecules at a rate which is unique to their previous growth condition [7, 8], therefore, the culture history of AOB may be another factor determining not only the rate of NH3 utilization [9] but also the rate of EE2 cometabolism.
For all of the reasons above, this study aims to determine effect of culture history on EE2 cometabolism by autotrophic AOB. AOB was enriched in mixed culture at different growth conditions. Effect of growth substrate concentration on bacterial cometabolism was also investigated. Although the past growth condition could potentially affect the bacterial cometabolism, but influence of the culture history of AOB on its EE2 cometabolism has never been investigated.
2. Materials and Methods
2.1. Chemicals
EE2 (17α-ethinylestradiol) was purchased from Sigma-Aldrich Corporation (St. Louis, MO, USA). The stock solution of EE2 was prepared to 500 mg/L in methanol. Other chemicals were purchased from Carlo Erba Reagents (Val de Reuil, France). All chemicals used in this study are analytical reagent grade.
2.2. Enrichment Media for AOB
Enrichment media, which consist of synthetic wastewater plus NH3 at various concentrations, was used for enrichment of AOB. Synthetic wastewater A was prepared by dissolving NaCl (1 g), MgCl2•6H2O (0.4 g), CaCl2•2H2O (0.1 g), KH2PO4 (0.2 g), and KCl (0.5 g) into one liter of de-ionized (DI) water and then adding one milliliter (mL) of each mineral solutions including the solution of non-chelated trace element, selenite-tungstate, vitamin mixture, thiamine, vitamin B12 [10]. Synthetic wastewater A was added with 0.34 g of (NH4)2SO4 per liter to achieve final NH3 concentration of 70 mg N/L. Synthetic wastewater B, which contained Na2HPO4 (4.05 g), K2HPO4 (2.1 g), MgSO4 • 7H2O (0.05 g), CaCl2 • 2H2O (0.01 g), and FeSO4 • 7H2O (0.09 g) in one liter of DI water [11], was added with 0.68 and 2.02 g of (NH4)2SO4 per liter to make up final NH3 concentrations of 140 and 420 mg N/L, respectively. Synthetic wastewater A and B were further used to prepare medium for batch degradation test.
2.3. Enriched Reactors Setup and Operation
Three vessels with an effective volume of 6 liters were identically set up as continuous flow enriched reactors without cell recycle. For each reactor, air was supplied from air blower connecting with a small stone diffuser to provide oxygen and mix the mixed liquor completely. The concentration of dissolved oxygen (DO) in bulk solution was maintained between 7–8 mgO2/L by adjusting air flow rate with a small needle valve. pH in mixed liquor was controlled at 7.5 ± 0.2 by a pH controller (Liquitron DP 5000, LMI Milton Roy, USA) equipped with a pH probe (Orion 9156DJWP, Thermo scientific, UK) as well as a solution of 0.5 N HCl and 0.5 N NaOH. Reactors were operated at room temperature between 23–25°C. Mixed liquor taken from full scale activated sludge process was used as inoculum to start up the reactors. Enriched reactors were operated differently in order to enrich AOB which belongs to different growth characteristics. Enrichment medium containing NH3 was continuously fed into each one of three reactors at different NH3 concentrations to enrich the AOB population which belongs to different rates of NH3 utilization. The reactors named ‘70NS’, ‘140NS’ and ‘420NS’ were fed with NH3 of 70, 140 and 420 mg N/L and controlled at sludge age of 4, 3, and 4 d, respectively. For each gram of added NH3, 0.068 grams of HCO3− was put into an enrichment medium to provide carbon source for AOB growth.
2.4. Batch Degradation Test
2.4.1. Reactor setup and operation
Several 25 mL small Plexiglas® vessels, which were initially filled with synthetic wastewater, were used as batch reactors. Sludge taken from enriched reactors during steady operation were washed and inoculated into the batch reactors. Synthetic wastewater was later filled into batch reactors until final sludge concentration of 300 mg mixed liquor suspended solid (MLSS)/L was reached. Then, a predetermined volume of the stock EE2 solution was added and fully mixed in order to achieve final EE2 concentration of 3 mg/L in the reactors. NH3 was later added into the reactors in a similar manner with that of EE2. Synthetic wastewater A was used particularly for tests with having initial NH3 levels of 28 and 70 mg N/L while for tests belongs to the initial NH3 concentration of 140 and 420 mg N/L, wastewater B was used instead. Air and mixing was provided to mixed liquor by shaking in an orbital shaker at 250 rpm. The reactor was incubated at room temperature. While running the tests, pH in bulk solution was maintained within a range of 8.0 ± 0.2 by adding HEPES buffer solution to a final concentration of 30 g/L.
2.4.2. Test procedure
Sludge containing the AOB population which belongs to different the NH3 utilization rates was taken from each one of three enriched reactors during steady operation and then, brought into batch degradation tests. Various initial concentrations of NH3 were used in the tests in order to determine effect of growth substrate concentration on the rates of NH3 oxidation and EE2 degradation. Loss due to abiotic sorption and volatilization of NH3 and EE2 were observed through batch tests with heat-killed cell and without cell. EE2 biodegradability by heterotrophic microorganism in enriched sludge was investigated in the presence of 10 mg/L allylthiourea (ATU) for the suppression of the AMO usage by AOB [12]. Furthermore, 1,000 mg/L chloramphenicol was also added into the reactors in order to evaluate the NH3 and EE2 biodegradability remained in the harvested sludge under the suppression of AMO-induced de novo synthesis [13]. All tests were carried out in triplicate except the test with heat-treated cells and with no cells were done in duplicate. For all tests, samples were periodically collected and analyzed for the concentration of EE2 and inorganic nitrogen species in bulk solution for the whole two weeks of the test period.
2.5. Chemical Analysis
Mixed liquor samples were collected from enriched and batch reactors and then, filtered through 0.45 μm Polytetrafluoroethylene membrane filter. Filtrate were analyzed for EE2 and inorganic nitrogen species (NH3, NO2−, NO3−) while filtrated were measured for MLSS and mixed liquor volatile suspended solid by gravimetric method. EE2 was analyzed by a C18 reversed-phase high-performance liquid chromatography with ultraviolet detector (Shimadzu Series LC-10ADvp) using a 60:40 acetonitrile:H2O as mobile phase. NH3 was measured by gas-sensitive combination electrode (Cole Palmer, Co.) and salicylated-colorimetric method with UV visible spectrophotometer (Thermo Electron Corporation, Hexious α, Cambridge, UK) at 640 nm [14]. NO2− was colorimetrically analyzed with UV visible spectrophotometer at 543 nm while NO3− was analyzed by UV screening method at 220 and 275 nm [14]. pH, DO concentration and temperature in mixed liquor were monitored during sludge enrichment and degradation test. pH value was determined by pH probe (250A+, Thermo Orion Inc., Germany). The concentration of DO was determined by membrane sensing electrode (DO 083005D probe with 850A meter, Thermo Orion Inc., Germany). Temperature was measured by a laboratory glass thermometer.
2.6. Analysis of Ammonia-oxidizing Bacterial Communities
2.6.1. Sample preparation and DNA extraction
Sludge samples (2 mg dry sludge/L) from two enriched reactors including ‘70NS’ and ‘420NS’ were concentrated via centrifugation at 14,000 rpm for 10 min and stored at −20°C. Genomic DNA was extracted from each samples (2 mg dry sludge) using Fast-DNA SPIN kits for soil (QBiogenes, USA) following to the manufacturer’s instruction. The extracted DNA was verified using 1.5% agarose gel electrophoresis (Bio-Rad, Spain).
2.6.2. Polymerase chain reaction (PCR), cloning and sequencing
Primers amoA 1F and 2R [15] (forward 5-GGGGTTTCTACT GGTGGT-3; reverse 5-CCCCTCKGSAAAGCCTTCTTC-3) were used for PCR amplification. Bacterial amoA gene fragments were amplified using a Takara polymerase (Takara Bio Inc., Japan) in a thermal cycler (Biorad Laboratories, USA) for 10 min at 95°C, followed by 35 cycles of 60 s at 95°C, 60 s at 56°C, and 30 s at 72°C, followed by 15 min of final extension at 78°C. Later, the amplified PCR products were purified by gel electrophoresis using a NucleoSpin Extract II Kit (Clontech Laboratories Inc., USA). Then, the purified PCR products were cloned using the pGEM-T Easy vector system (Promega, USA). Later, five and four clones of ‘70NS’ and ‘420NS’ samples were randomly chosen for sequencing at Macrogen Inc., Korea, respectively.
3. Results and Discussion
3.1. Enriched Reactors Performance
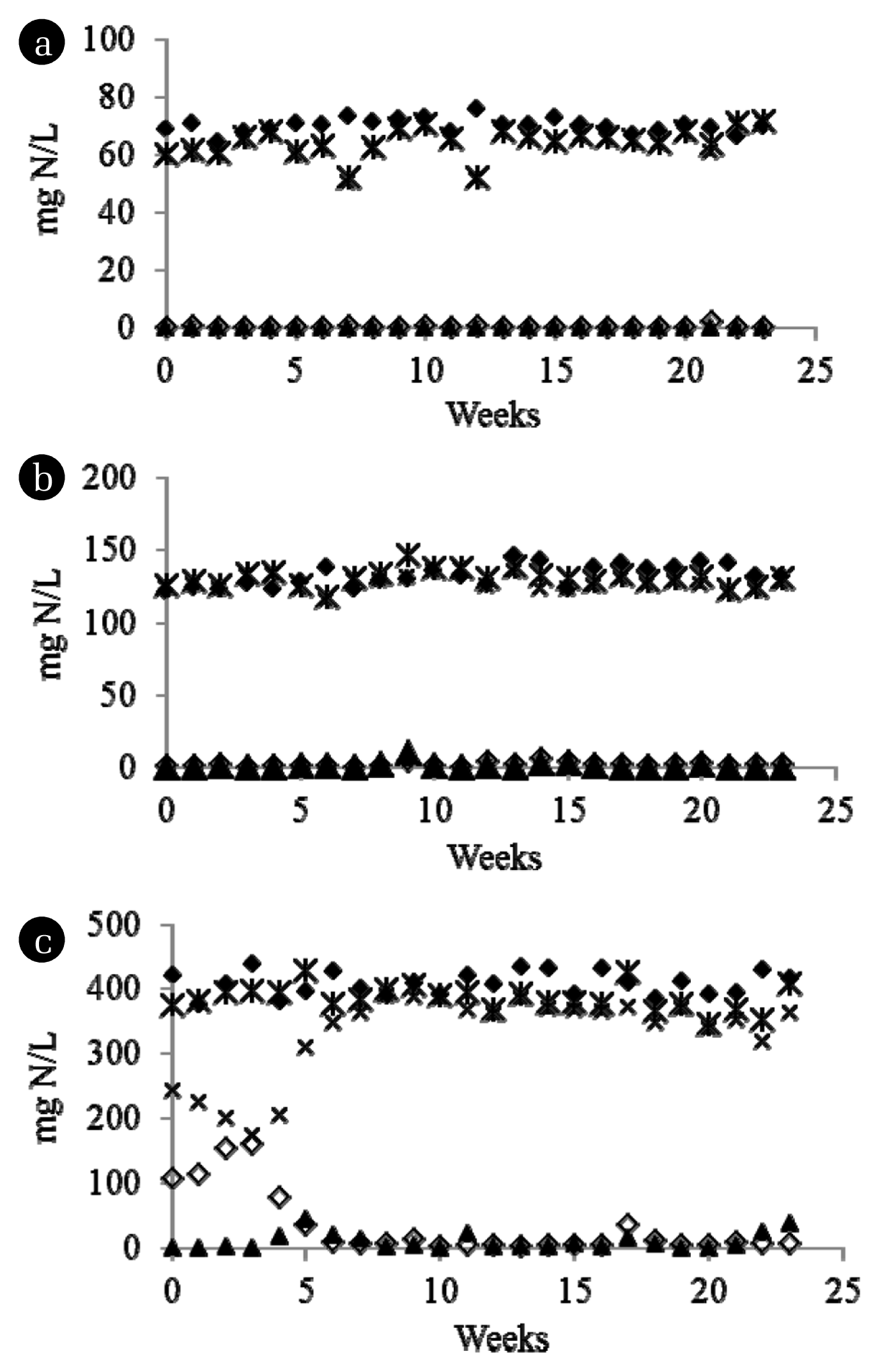
Table 1 and Fig. 1 showed long-term performance of all three enriched reactors. All reactors showed steady performance in terms of NH3, NO2− and NO3− effluent concentrations after five weeks of operation. At steady state condition, NH3 removal efficiencies for reactors fed with NH3 of 70 mg N/L and 140 mg N/L were more than 99%. But for 420 mg N/L NH3-fed reactor, a slight less efficiency for NH3 removal (96.23%) was observed due to receiving higher NH3 load compared to other reactors. In addition, an increase in NH3 load led to stimulate specific NH3 oxidation rate. No NO2− accumulated in any reactors indicating NH3 oxidation is the rate determining step for nitrification.
3.2. Ammonia-oxidizing Bacterial Communities in Enriched Sludge
Sludge ‘70NS’ and ‘420NS’ were brought to further analyze for ammonia-oxidizing bacterial communities using PCR-cloning- sequencing technique. Results showed that AOB belongs to Nitrosomonas oligotropha (N. oligotropha) cluster dominated (4 of 5 clones) in reactor fed with NH3 of 70 mg N/L at which the NH3 concentration were less than 1 mg N/L in effluent (Table 1). AOB belongs to N. oligotropha cluster is known as a high NH3 affinity cluster as indicated by low Monod half-saturated coefficient (Ks: 0.32–3.16 mg N/L [16, 17]). However, reactor fed with 420 mg N/L NH3, which its effluent contained 15.82 mg N/L NH3, was dominated (all 4 clones) by AOB belongs to Nitrosomonas europaea-Nitrosoccus mobilis (N. europaea-N. mobilis) cluster. AOB belongs to N. europaea-N. mobilis cluster has much lower NH3 affinity than that belongs to N. oligotropha cluster as indicated by much higher Ks (1–100 mg N/L, [16–19]). These results corresponded with the finding reported in Limpiyakorn et al. [20]. Limpiyakorn et al. [20] showed that N. europaea-N. mobilis cluster dominated in chemostats fed with > 140 mg N/L NH3 while for the reactor fed with ≤ 140 mg N/L NH3, the dominant AOB belonged to N. oligotropha cluster.
3.3. Batch NH3 Oxidation
3.3.1. Effect of culture history
In order to investigate effect of continuous culture history on batch substrate utilization, three enriched sludge being acclimated with various loads of the growth substrate (‘70NS’, ‘140NS’, and ‘420NS’) were brought to determine the rate of substrate utilization under batch condition. Results showed that continuous culture history of enriched sludge strongly influence on batch NH3 oxidation rate (Table 2). For any given values of initial NH3 concentration, higher NH3 oxidation rate was found in sludge previously being acclimated with higher load of NH3.
This is likely because continuous cultivation of mixed sludge at higher NH3 load leading to the selection for a particular AOB having higher NH3 utilization rate. Generally, the dominant bacterial population mainly determined the rate of substrate utilization and bacterial growth of mixed culture. Sludge dominated by AOB with N. europaea-N. mobilis cluster (420NS) showed higher batch NH3 utilization rate as compared with that of dominated by N. oligotropha AOB cluster (70NS). For all results shown above, an intrinsic NH3 oxidation rate clearly depended on the physiology of dominant AOB population in enriched nitrifying sludge.
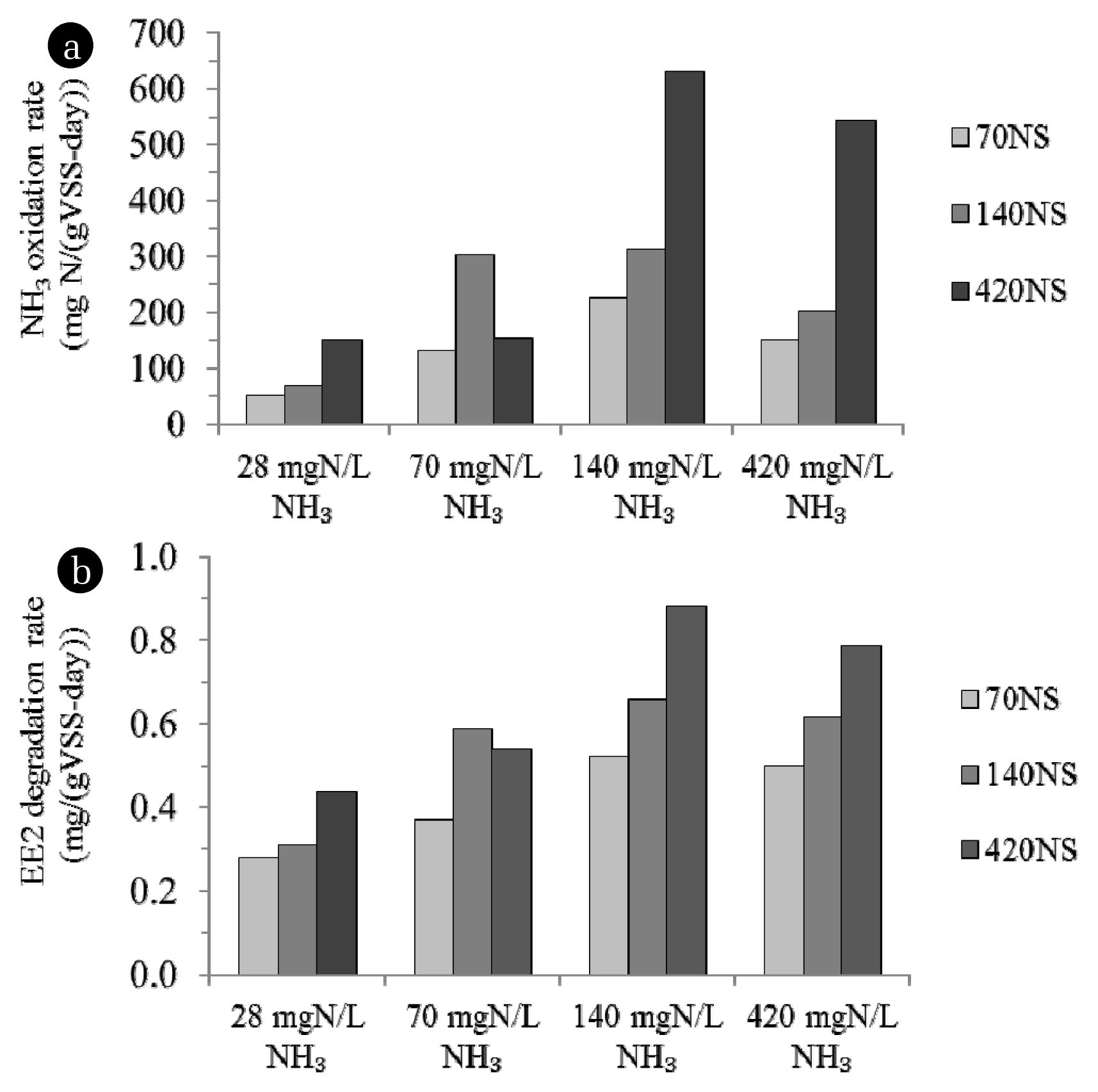
3.3.2. Effect of initial NH3 concentration
As a result, NH3 oxidation was limited by substrate availability. Larger specific NH3 oxidation rate was observed at higher initial NH3 (Fig. 2). However, maximum batch NH3 oxidation rate of enriched sludge was attained under initial NH3 concentration which had been previously used for feeding during sludge enrichment. Enriched sludge in reactors fed with 70 mg N/L NH3 (70NS) achieved its maximum NH3 oxidation rate at 70 mg N/L while at least 140 mg N/L was needed by sludge taken from the 140 and 420 mg N/L NH3-fed reactors (140NS and 420NS). In other words, N. oligotropha-dominating sludge (70NS) achieved its maximum NH3 oxidation rate at lower NH3 concentration than that of sludge dominated with N. europaea-N. mobilis AOB cluster (420NS). Higher NH3 affinity of the 70NS as compared to that of the 420NS because it was dominated by the N. oligotropha cluster which has higher NH3 affinity than the N. europaea-N. mobilis cluster.
Moreover, at initial NH3 concentration of 420 mg N/L, slight lower NH3 was oxidized as a result of substrate inhibition. However, sludge dominated with N. oligotropha AOB cluster (70NS) were apparently affected far less by the substrate inhibition than N. europaea-N. mobilis-dominating sludge indicating higher tolerability to adverse condition in slow-growing AOB. This emphasized an importance of past cultivation condition of sludge on its rate of substrate utilization in batch reactor.
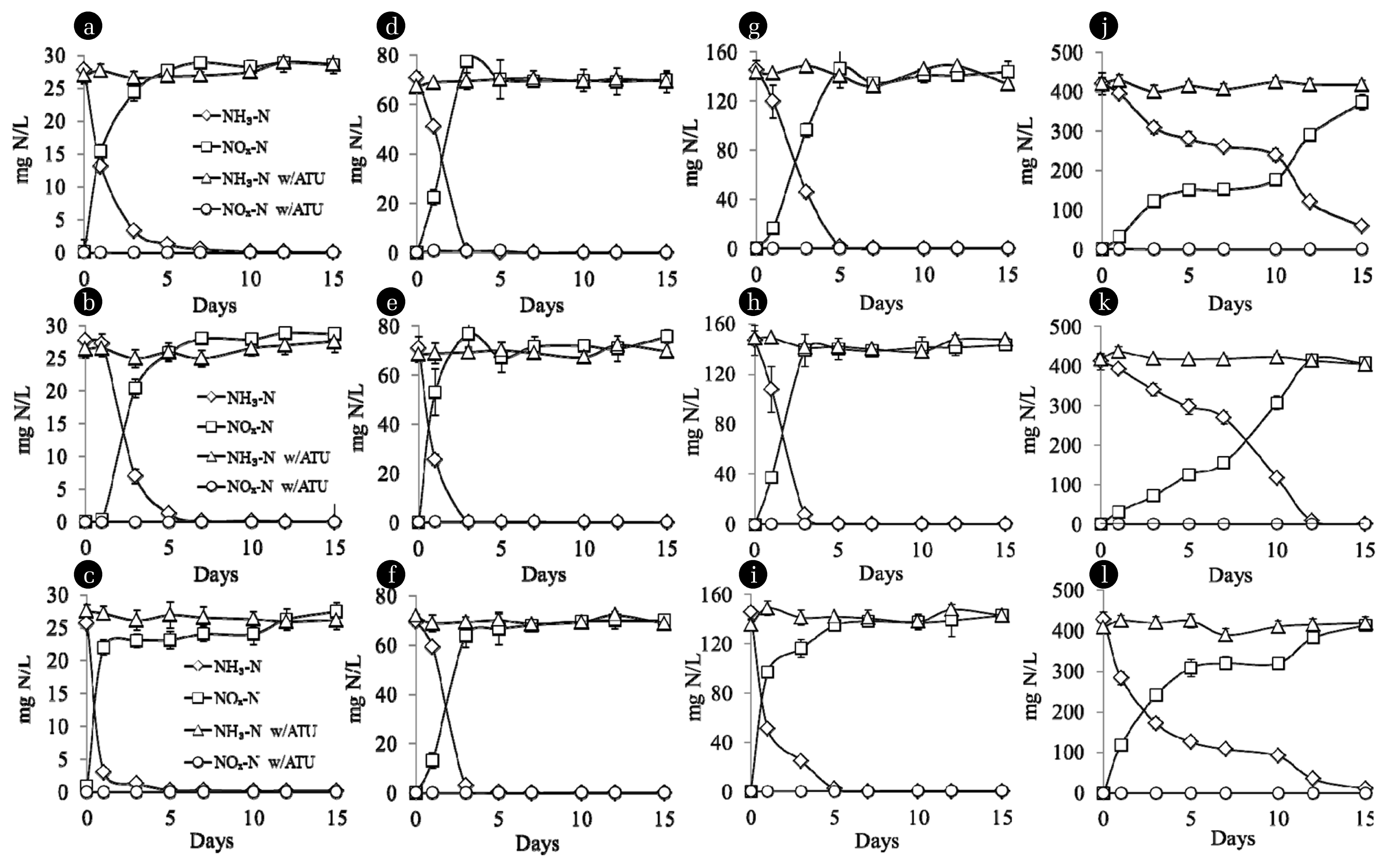
3.4. Batch EE2 Degradation
EE2 biodegradation by sludge enriched with different loads of NH3 was investigated to observe effect of culture history on the AMO cometabolism. As shown in Fig. 2 and 3, NH3 and EE2 degradation was observed only in the absence of ATU. Furthermore, there was not any EE2 degradation in batch with heat-treated cells or no cell (data not shown) indicating the loss of EE2 due to abiotic processes (ie. sorption and volatilization) was negligible in our study. This indicates that AMO cometabolism by AOB is main degradation pathway of EE2 rather than heterotrophic and abiotic degradation that is correspondence to several studies [12, 21–25]. Moreover, EE2 and NH3 degradation in the presence of AMO-induced inhibitor was not found (data not shown) indicating residual AMO utilization by harvested cells in order to complete both substrate degradation was negligible.
EE2 biotransformation clearly depended on NH3 oxidation. During EE2 cometabolism, EE2 was degraded by using a reducing power from NH3 oxidation. Therefore, more NH3 was oxidized, higher EE2 was degraded via the cometabolism of AMO. Moreover, not only initial NH3 concentration largely affected on NH3 oxidation rate but on EE2 biotransformation rate as well. Higher available NH3 improved both the rates of NH3 and EE2 removals indicating a reductant-limiting condition for AMO metabolism and cometabolism. While a reductant-inhibiting condition decreased NH3 oxidation and EE2 degradation at initial NH3 of 420 mg N/L. The results shown here is in good agreement with previous studies [22–24] which found strong positive correlation between initial concentration of NH3 and batch EE2 biotransformation rate by nitrifying sludge.
In addition, after 5 d of sludge incubation with initial NH3 of 28–140 mg N/L, all NH3 were utilized, but substantial biotransformation of EE2 was still occurring even in the absence of NH3. This was consistent with the results obtained by a number of studies [22, 26–28] which revealed the capability of nitrifying sludge to cometabolize EE2 in NH3-free environment. This phenomenon is likely to be a result of utilizing AMO accumulated within the AOB cell for degrading EE2 during NH3-limiting condition.
Moreover, EE2 degradation of enriched sludge was affected by not only initial NH3 level but also the enrichment condition. Higher EE2 biotransformation rate was achieved by sludge previously being enriched with higher NH3 load (Fig. 4). As mentioned earlier, the enrichment of nitrifying bacteria at higher NH3 load may promote specific AOB having higher utilization rate of NH3 over those having the lower one. The results show that AOB population in enriched nitrifying sludge affected on the EE2 degradation in the same manner as NH3 oxidation. Higher EE2 degradation was found in sludge dominated by higher-growth-AOB than lower-growth- AOB-dominating sludge. EE2 cometabolic rate of sludge dominated with the N. europaea-N. mobilis AOB cluster were approximately 1.6–1.7 (420NS) times higher than that belongs to the N. oligotropha AOB cluster (70NS). Greater EE2 cometabolic rate of higher-growth-AOB may be caused by more electron available for cometabolism.
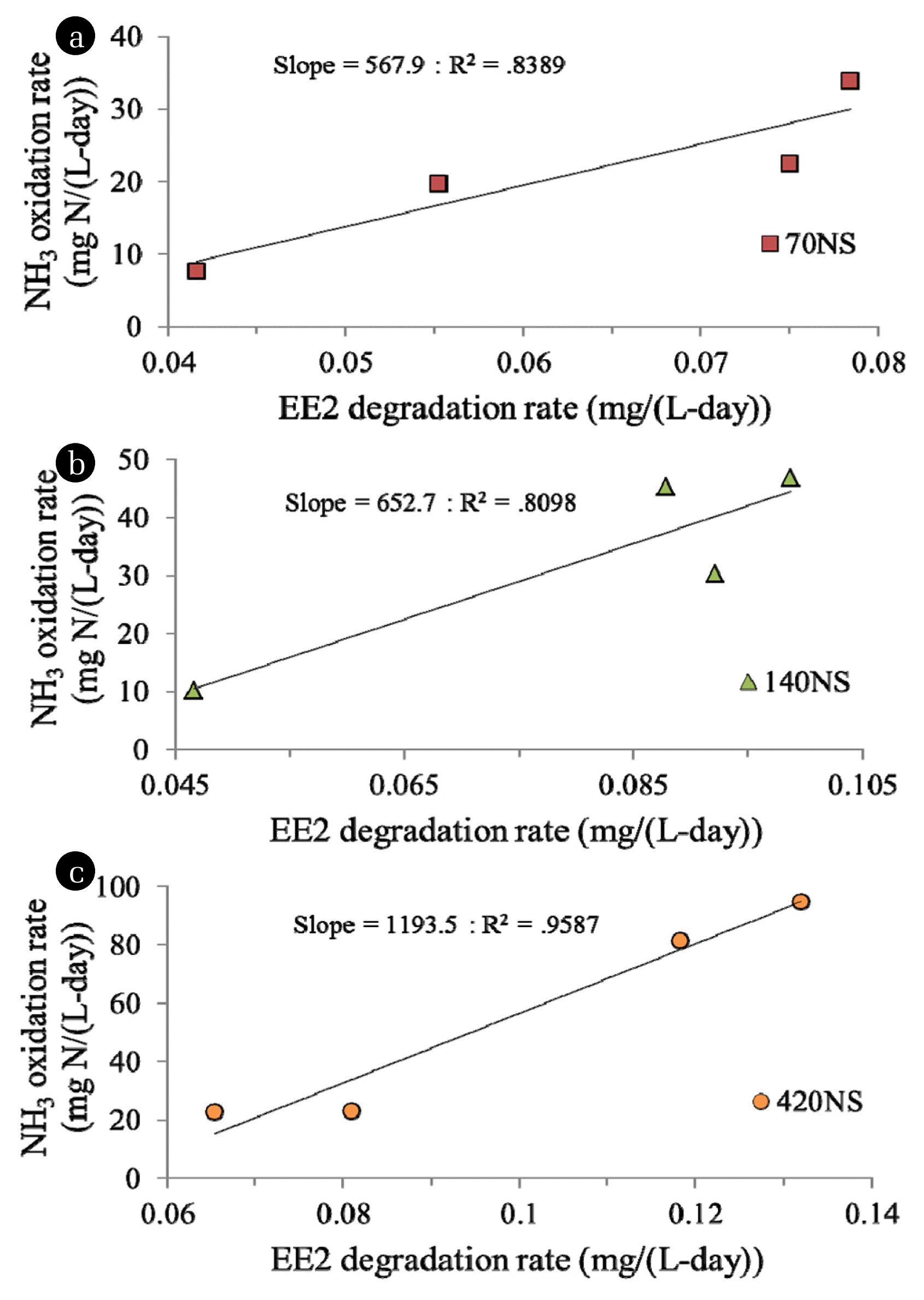
Furthermore, the correlation between NH3 and EE2 degradation rates of sludge was found strongest in the sludge being previously cultivated with highest NH3 load (Fig. 5). It can be implied that more electron provided by NH3 oxidation is needed by sludge being previously cultivated with the highest load of NH3 to cometabolize a certain amount of EE2. Our finding firstly reveals the importance of past AOB growth condition regarding its determination on simultaneous N and EE2 removals. The rates of EE2 biotransformation found in this study were compared with those reported in previous studies (Table 2). Higher rates of EE2 degradation obtained in this study as compared to other studies is likely because much higher initial concentration of EE2 was used.
4. Conclusions
This research firstly determined the importance of culture history of nitrifying sludge on simultaneous NH3 and EE2 removals in wastewater. Higher available NH3 improved both the rates of NH3 and EE2 removals indicating a reductant-limiting condition for AMO metabolism and cometabolism. Sludge being acclimated with higher NH3 load as well as the one dominated with higher-growing-AOB showed higher rate of EE2 cometabolism due to its ability to provide more reducing power from NH3 oxidation. The NH3 and EE2 degradation rates were found having a stronger correlation particularly in sludge being previously cultivated with higher NH3 load.